More than 25.000 natural orchid species are known, and insects pollinate the vast majority. Some species are pollinated by different insect species, but many are so specialized that only one particular insect species is able to pollinate the orchid. This means co-evolution has happened between orchid and insect. The system secures an effective cross-pollination but increases the risk that both species become extinct if life condition for just one of the species changes in an unfortunate direction. This page illustrates a number of the most refined cases of interaction between flower and pollinator, but first it is explained how the orchid flower is constructed.
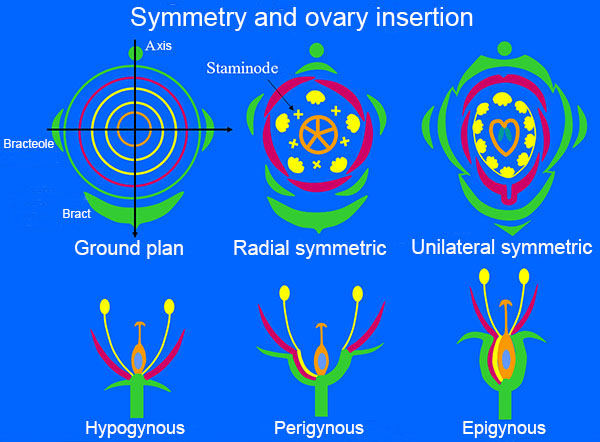
A flower consists of a number of leaf-whorls, innermost carpels, and outermost sepals. The number of leaves in each whorl varies and often there is only one whorl of stamens, although, two are shown on the basic diagram (ground plan). Reduction of leaf number in a whorl or fusion of leaves occurs in many species. Reduced non-functional stamens are called staminodes. The insertion of the flowers with relation to the ovary is shown lower left. The orchids have epigyneous flowers (the ovary is inferior ) i.e. the tepals (sepals + petals) and stamens are fused with the carpels almost to the tip of the carpels. The orchid flower is zygomorf, i.e. there is only one plane of symmetry just as in the pea family. The starting point corresponds to a three-merous liliaceous flower and therefore orchids and lilies were by some botanists placed in the order Liliales. Many orchids have a spur from the abaxial petal and while the bud develops, the ovary and flower stalk wind, so the spur in the open flower points away from the axis of the inflorescence and the flower is said to be resupinate.
It is easy to see the resupination in Vanda, since the ovary is furrowed corresponding to the dorsal and ventral suture of the three carpels. However, the ovary can regulate the winding if the inflorescence is skew or bowed so the flower as a general rule is oriented with the lip at the bottom, i.e. in the direction of gravity. This is probably the most appropriate position in relation to the pollinator, which normally uses the lip as a landing site.
The variation in sepals and petals is shown above. In Vanilla imperialis (upper left) they are almost equal except for the lip. This is probably the most common. In Masdevallia tovarensis (lower left) the sepals dominate, while the petals and the lip are vanishingly small. In Phragmipedium phillippinense the two lateral petals are extremely long, narrow and winded while the lip is shaped as a slipper.
Examples of variation in the lip. The Phalaenopsis hybrid (left) has a large lip with wings. Coelogyne lawrenciana (middle) has a similar shaped lip but equipped with papillae. Renanthera imschootiana (right) has almost no lip.
In Stanhopea oculata (left) the lip is almost monstrous with an eye spot and a fork-shaped ending. Aerangis biloba (right) exemplifies the lip can be equipped with a spur containing nectar. In some species, the spur is just a small bagging, in others it can be more than 30 cm long. Other tepals may also be equipped with a spur.
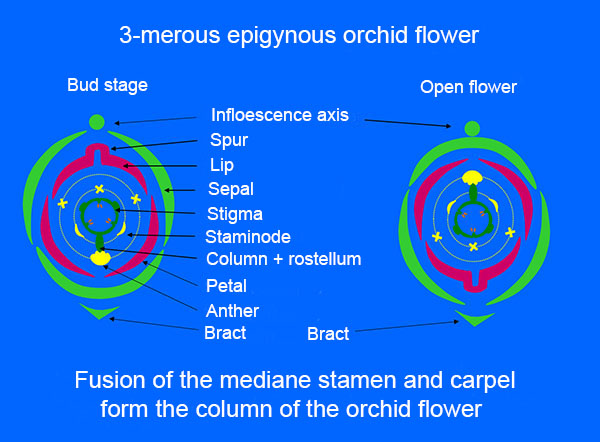
The fusion between the only developed stamen and the nearest carpel to form the so-called column (gynostemium) is probably the most special thing about orchid flowers. Simultaneously, the stigma of the two lateral carpels are also fused and the mediane stigma forms an outgrowth called rostellum. It is a beak between stigma and the anther to prevent self-pollination. The tip of the rostellum is provided with an adhesive disc (viscidium) which plays a role when the pollinia from another flower are to be picked up. Orchid pollen stick together in pollinia, which on the right illustration are isolated from the anther.
Zygopetalum mackay. At left, the arrow points to the anther and the rostellum. The pollinia are exposed in the middle photo. The stigma is placed in a cavity on the ventral side of the column. The surface of the stigma produces a sugary fluid (glistens on the pictures), where the pollinia are caught and germinate. At right the flower is pollinated, the surface of the stigma has dried, and it has turned brownish. The anther is empty.
The stigma can be three-partite or bipartite, but most often the two stigmas are fused as in Rossioglossum grande (above). The tip of the rostellum is almost black in this case.
The diagram shows different methods of pollination in flowering plants. Insects pollinate by far most of the orchids, but birds are also on the list. Self-pollination may occur, and it is not uncommon that self-pollination happens as a last resort. If a flower is not pollinated by an insect, the pollinia may fall out and hit the stigma shortly before the flower wilts and thereby self-pollination occurs. Ophrys apifera is always self-pollinated and hence by an evolutionary end station. A few years ago, it was found near Vejle as a new species for Denmark. Photo: Woodward, Flickr.
In orchids with female and male plants as in Cycnoches chlorochilon self-pollination is of course impossible. In this species the two genders look much alike, but in other species the difference may be considerable. It is female at left and male at right. Notice the long curved column. About pollination in Cycnoches see later. Photo of female flower: J Pestana, Flickr.
The bisexual flower has streamlined insect pollination since the same insect is able to both collect and deliver pollen at the same visit. However, no insect pollination unless there is something to lure the insect. This can be food, resting place, heat, hiding place, or a place for oviposition. At left a bumble bee has filled its pollen basket on the hind legs during a visit in Cosmea but it does not work with orchids. Orhids cannot afford the pollinia becoming eaten. All pollen are needed because orchids produce a vast number of small seeds. 3,770,000 seeds have been counted per capsule in Cycnoches chlorochilum. Therefore, orchids must have something else to offer and that is first and foremost nectar. However, it may give the problem that orchids using pollen-collecting insects as pollinators indirectly become dependent on the availability of other plants that can deliver pollen. At right Platanthera bifolia with long spurs filled with nectar.
Despite nectar being an important luring agent, one does not reckon that more than two thirds of all orchids produce nectar. The remaining species offer other food and fragrances. E.g. at least 50 of Ecuadors Maxillaria species offer a light powder on the lip produced by glandular hairs. The powder replaces pollen and lure pollen collectors. The powder consists of ligated cells containing starch and oils. At left Maxillaria grandiflora (Photo: Mauro Rosim, Flickr). Callus on the lip may consist of hairs (trichomes) or papillae secreting oils. Many insects chew on the hairs and thereby obtain a food reward for visiting the flower. At right, Coelogyne cristata.
When flowers are less than 1 cm in size, insects easily overlook them. Therefore, small flowers are collected in inflorescences, which can be seen clearly from a larger distance. In Oncidium truliferum certain bees are attracted by the inflorescence because they perceive it as a swarm of bees.
The recognizability of the flower also plays an important role in attracting a particular insect species. This is about shape, size, colors, UV-patterns, and scent. Insects recognize more easily symmetrical than asymmetrical shapes. The zygomorphic flower, i.e. with one plane of symmetry is attractive to insects. The picture shows Calypso bilbosa, which is circumpolar in the coniferous forest belt
Some orchids make use of mimicry to lure insects, i.e. the flowers look like a female insect and therefore the male attempts to copulate. Ophrys insectifera is pollinated by male Argogorytes mystaceus and A. fargei (family: Crabronidae). The male performs a pseudo-copulation. The males develop two weeks before the females and hence they have no one to mate with. The flower imitates in a unique way the look of the female and furthermore it produces a stronger scent of sex hormone than the female. Notice the two lateral tepals in the inner whorl (petals) form ’antennae’. It has been shown the flower shape is not particularly important in luring the males. If a part of the lip is cut off, the flower is visited just as often as long as the scent glands are intact. There is no nectar in the flower and therefore the wasps visit the flower only for copulation. This makes the orchid dependent on other plants being available as food source for the male insects. When the female wasps develop, the flowering of the Ophrys has culminated. When the male has visited an Ophrys flower, it scents as a female and the copulation pattern may be a very different one.
Another kind of mimicry which is also pure cheating and deception is seen in the Australian orchid Thelymitra antennifera. It does not produce nectar but it looks like a lily, which has nectar and therefore it is visited by nectar seeking insects. Photo: P Bouchard, Flickr.
Ascocenda Thai Lemon
Dendrobium lawesii
Dendrobium victoria- reginae
Coelogyne speciosa
In the orchid family every possible color is realized in the flowers. The many colors and combination of colors reflect a very large number of different insects acting as pollinators but as mentioned, it is often only a single insect species that is able to pollinate one or a few orchid species. – In this connection, it has to be mentioned that there is a considerable difference in how insects perceive colors and their color perception is very different from ours. E.g. many bees are unable to see red, while certain butterflies have no problems with red. Generally, the insect color vision is dislocated relatively to ours, and to bees the green leaves are greyish-black. Thereby the flowers will appear luminous on a dark background.
Scent is another important factor in order to attract pollinators to orchids. Many people have a perception that orchids have no scent. It is probably because Phalaenopsis hybrids are common houseplants and they have no scent. A fragrant Zygopetalum mackayi from Brazil is shown above at left. – At right, an Elephant Hawk-moth (Deilephila elpenor) protrudes the trunk (Photo: J Mangelsdorf, Flickr). Experiments have shown the moth protrudes its trunk in wind tunnels when flower scents are served, and the Hawkmoth Sphinx ligustri is able to orientate alone by scent without using vision. In the Amazonas rainforest in Ecuador orchid bees (Euglossa sp.) pollinate Chondrorhyncha caudata. The male bee is attracted by the scent glands of the orchid, which the bees collect on the hind legs in order to seduce the females. When the bee leaves the flower, it often happens with pollinia attached to the head. – Birds, on the other hand, have no sense of smell. In the mountains, birds often replace insects as pollinators.
Nectar is an obvious lure. It is most often produced in a spur on the lip as seen in Platanthera bifolia (left) and Aerangis thompsonii (middle), but nectaria may also be placed at the base of petals and sepals or on the column. The level of nectar can be sensed at the bottom of the spur of P. bifolia (see photo). In A. thompsonii the lip is bend backwards since it is pollinated by nocturnal moths. These insects stand in the air and swirl its wings and they do not need a landing place. Nectaries may also be extrafloral and placed on vegetative parts as shown at right where a droplet of nectar is visible at the base of each flower stalk in Cattleya. The function of these nectaries is to prevent crawling animal from reaching the flower.
There are still some methods for luring insects to flowers, but they are of no importance to orchids. Some species use advertising flowers. It is barren flowers with the purpose to make fertile flowers more visible. Examples are the peripheral flowers on composites (Compositae/Asteraceae) and Hortensia (Hydrangea) shown at left. At right, it is Lysichitonamericanus (Araceae), where the yellow spathe makes the inflorescence more visible. This species has further the sophistication that the flower spike produces heat, which attracts insects, in this case small flies.
A successful pollination is also dependent on how long the flower is open and durable. A self-pollinated flower need not open at all, but in general, orchids excel by being durable over many days and even months. It may be related to the fact that there is often a long distance between two individuals of the same species. The dependency of a particular insect species becomes less critical when the flower has a long opening time. Stanhopea species only last 4-5 days but they compensate by having very large flowers with a strong scent. Cattleya flowers last 5-8 weeks. Phalaenopsis amabilis and its hybrids (at left ’Alice Gloria’) may last for up to four months, but that is short time compared to Grammatophyllum multiflorum from SE-Aasia where the flowers are durable three quarters of a year. As a contrast it may be mentioned that bat pollinated cacti such as Cereus jamacaru (right) only last one night.
It costs a lot in energy to keep the flowers fresh over long periods and therefore it is an advantage, if the flower can contribute to cover the energy consumption through its own photosynthesis. Many orchid flowers are greenish due to the presence of chlorophyll. In a single Cymbidium species it has been proven that the flower performs photosynthesis. This, no doubt, is also the case in Coelogyne pandurata (Malaya-Borneo, left photo). – Both C.pandurate and Dendrobium margaritaceum (right) illustrate that a place is needed where the insect can land when visiting the flower. In a zygomorphic flower, it is always the lip, which functions as a landing site. By sitting here the insect is immediately correctly orientated both in relation to searching for food and for pollination. The landing site has both red and yellow ‘lights’ as in another airport. In other species colored stripes on the lip function as directories. See e.g. the already shown photos of Calypso bulbosa, Coelogyne cristata, and Zygopetalum mackayi.
Plenty of pollen and large stigmas are characteristic for wind-pollinated flowers. Orchids must also produce plenty of pollen due to the many ovules needing fertilization. In the majority of species the pollen is transferred on once glued together as pollinia as illustrated above for Embreea rodigasiana (Columbia-Eucuador) including the viscidium which has been attached to the pollinator. The stigma is relatively large (brownish in Rossioglossum grande at right) maybe to ensure greater accuracy in the pollinia transfer so also skewly placed pollinia can be captured. The position of pollinia and stigmas are important to maximize the precision during pollen transfer and simultaneously minimize energy consumption. The fusion between the functional stamen and the carpel in the column serves this purpose specifically, and at the same time, it is the foundation for the strong specialization of the flower that only a single insect species or sometimes even a single gender of the insect is able to perform the pollination. Such an adaptation has taken millions of years to develop and it has happened by a parallel specialization of plant and insect called co-evolution.
The pictures above show especially the design of the lip with upwardly bent wings limiting the pollinator’s freedom of movement. That means if the insect should be able to reach the food, it must enter the tube and there is only one way forward. There is also only one way back, when the pollinator backs out again the barb-shaped rostellum will adhere to the animal, and the viscidium will tear loose so the pollinia will follow and stick to the pollinator. The shown bumblebee is visiting Schomburgkia superbiens in Denmark and since it is not a natural pollinator it looks as if the pollinia are placed inappropriately too far back on the wings of the bumblebee. S. superbiens grows as an epiphyte in mountain forests of Central America up to 2,000 m.a.s.l.
The cooperation between orchids and the pollinating insects goes so far that many orchids signal when they are pollinated so it is not worth it to visit the flower since the nectar is taken and nectar production has stopped. The signals are diverse and may consist of color changes in parts of the flower, changes in scent, swellings of flower parts, hormone production, and more. – During wilting of the flower, some of the degradation products are withdrawn for reuse in the plant. The wilting is initiated by ethylene production, which starts when the anther hood and pollinia are removed unless the flower is pollinated. Investigations of such post-pollination phenomena in Phalaenopsis about 160 years ago led to the discovery of the cell nucleus and about 100 years ago to the discovery of the plant hormone IAA (indoleacetic acid), which is an auxin. Notice on the middle picture of Zygopetalum mackayi that the flower still has its own pollinia besides having received a pair from another flower. These pollinia appear shrunk because the pollen have germinated and grown down the column to the ovules. Simultaneously the column is swollen around the stigma. At right, the pollinated flower is shown above. The ovule has increased in thickness compared to the wilted non-pollinated flower below, and the petals are still green and able to contribute with photosynthesis.
Self-pollination occurs regularly in about 3% of all orchid species. Some are even apomicts i.e. they develop seeds without fertilization. In Denmark, self-pollination is known from several species of Cephalanthera. In this genus, the rostellum and viscidium are lacking and pollinia and stigma are poorly separated. Left photo shows C. damasonium, where the flowers almost do not open. At right, it is Liparis loeselii, which is self-pollinated by help of rain drops that knock the pollinia out of the anther thus they hit the curved lip so that the pollinia strike up against the stigma.
One assumes insect pollination has developed from wind-pollination. At left, a dust cloud of pollen is carried away from Juniperus communis by wind. Primitive wind-pollinated gymnosperms have just as our day’s conifers secreted a sticky substance around the mouth of the ovule (the micropyle) to catch the wind carried pollen. Beetles crawling on the plants have discovered a food source in the pollen and the sticky substance. They have then more regularly sought out the ovules and they could not avoid transferring pollen from plant to plant and thereby making pollination more effective. The right photo shows young female cones of Thuja occidentalis with liquid drops on some of the ovules. Thuja photo: Plant Image Library, Flickr.
Some Orchis species like the above shown O. militaris (left) and O. purpurea and many other Orchis and Dactylorhiza species are pollinated by solitary bees and bumble bees.
The fly orchid’s (Ophrys insectifera) interaction with male wasps is already discussed. South European species such as Ophrys speculum (Left photo: M Freeth, Flickr) and Ophrys tenthredinifera (middle) also use the similarity with insects to lure males of different bees. Ophrys lutea (right) is pollinated by a male bee of the genus Andrena but in this case the lip imitation of the female is oriented in opposite direction, i.e. the head is turned away from the center of the flower, the copulation takes place correspondingly, and the pollinia are placed on the abdomen of the male. Lower photo: D Attenborough 1995.
The lady slipper Cypripedium calceolus is pollinated by five different smal Andrena bees, which often end falling into the lip. There are translucent parts on the back wall (red circle) which shows the direction to the way out at the red arrow, which points at an anther. Cypripedium and the closely related Paphiopedilum differ from the majority of orchids by having two functional anthers, one on each side of the column, hence the bee will encounter one anther independently of which of the two ways out it chooses.
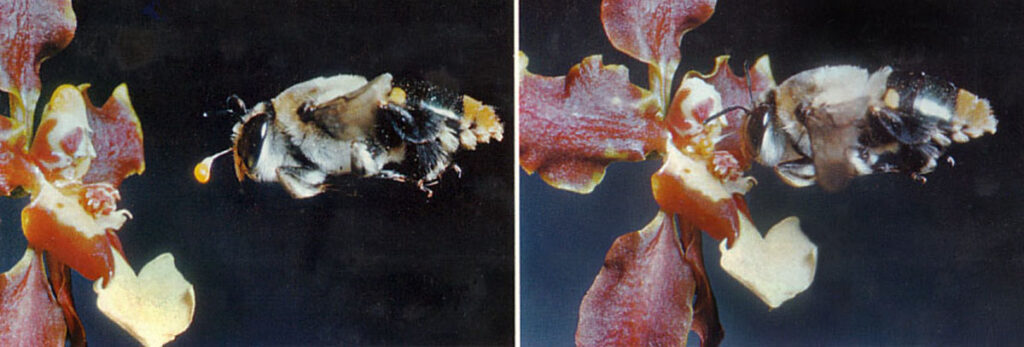
As earlier mentioned Cycnoches (Central- and South America) has unisexual flowers, which in some species are almost identical, while in others they are very different. Above the female C. chlorochilon is seen at left (Photo: J Pestana, Flickr) and the male is at right. The lip is turned upwards. Euglossine bees (tribe Euglossini, subfamily Apinae) are tropical bees where the males are attracted by certain scents, which have an anesthetic effect. The flowers rarely contain nectar. The male bee lands on the upper side of the lip and it then slides down the lower side where it must let go with its hind legs in order to reach the scent area at the tip of callus. The heavy abdomen swings down and hits the column. This releases the pollinia, which are under tension. The pollinia stick to the bee near the tip of the abdomen. The same thing happens in the female flower, where the pollinia are deposited on the stigma when the bee takes off by letting itself fall from the lip. – Cycnoches egertonianum is a species with large differences between the two genders. It is visited by the male Euglossa hemichlora. The lip has a hinge so the outer part swings down due to the weight of the bee and then the abdomen hits the anther or the stigma.
Stanhopea tigrina is likewise pollinated by male bees attracted by a strong scent, which is produced by a glandular tissue in the hollow lip. The middle lobe of the lip and the side lobes form a chute which the bee slides backwards downwards when it has been well anesthetized by the scent. The passage between the very long curved column and the lip is so narrow that the back of the thorax repels the rostellum and the pollinia become attached. If pollinia are brought by the male, they are deposited on the stigma. Since the flower is protandrous, self-pollination is not possible. The pollinia are thick, while the stigma is narrow and sunken until at maturity it swells.
The different design of the lip and column in Stanhopea is a good example that each orchid species uses one or a few related male bees for the pollination. The already shown Stanhopea cucullata has a smaller flower than S. tigrina but a broader funnel between lip and column adapted to a larger bee species than the one pollinating S. tigrina. What the function is of the eye spot is unknown to me. Above at left S. martiana from Mexico is shown and at right S. napoensis from Ecuador.
In Gongora the lip is turned upwards and it is shaped fairly complicated. When the male bee lands on the lip, it turns around so it hangs upside down under the lip, where it scrapes after fragrances. They are found at the base of the lip so the bee must move in between the forks, which will steer the bee when it loses foothold and it falls on its back down on the column, where the pollinia are then collected on the back. The left photo shows Gongora quinquenervis. It is a variable species found from Central America to Peru. The right photo is G. galeata from Mexico. It is pollinated in the same manner except that the male bee is another species.
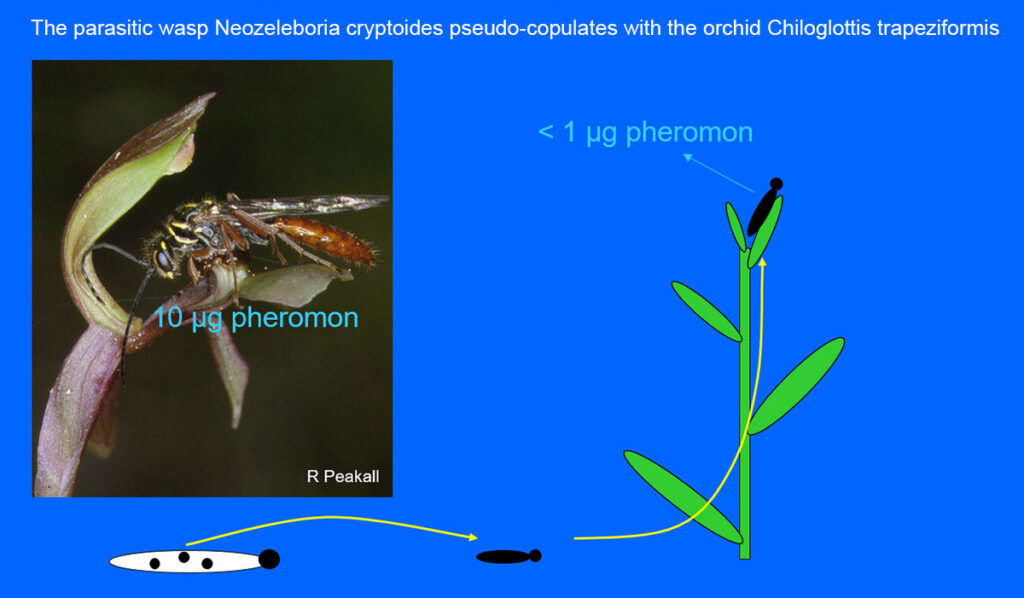
Several orchids emit female pheromones in order to lure certain insect males. This is already exemplified by the Fly orchid Ophrys insectifera. Some other examples from Australia follows. The wingless female of the parasitic wasp Neozeleboria cryptoides lays its eggs in insect larvae. When the female emerges from its pantry, she depends on the winged males to provide food. In order to obtain his attention she crawls up to the top of a plant and emits pheromones. When the male finds her, they copulate and then he carries the female from Eucalyptus tree to Eucalyptus tree where she feeds on secretes from aphids until the male finally delivers her to an insect for laying eggs. However, if the orchid Chiloglottis trapeziformis is nearby, things go wrong for the parasitic wasp because the orchid emits ten times as much female pheromone as the wasp female (10µg against 1µg), and since the black callus area of the lip imitates the female wasp, the male choses the orchid and tries to pseudo-copulate. Simultaneously, the male delivers pollinia if it brings some with it from another flower.
The parasitic wasp Neozeleboria cryptoides pseudo-copulates with the orchid Chiloglottis trapeziformis. The arrow in A points to the stigma. In B the pollinia is delivered. After visiting 4-5 flowers the male learns its mistake, therefore the orchid must constantly develop stronger attractive means so the next generation can also be pollinated. This is an example of stepwise co-eveolution. Photo: M L de Jager & R Peakall 2019, Ann. Bot. 123: 347–354.
Above three examples of Australian Hammer orchids. They are orchids where the lip has a hinge, which in particular parasitic wasps (thynnine wasps) can activate when they land on the lip. Then the lip and the wasp swing against the column where pollinia may be collected or delivered. Once again, it is pheromones and physical similarity with the female wasp that attracts the male wasp. Drakaea thynniphila is shown above left (Photo: P Bouchard, Flickr). Below left, it is Drakea livida (Photo: Jean & Fred, Flickr). The other four photos is D. glyptodon (Photo B Bohman et al. 2014, New Phytol 203,3). It is pollinated by the male thynnine wasp Zaspilothynnus trilobatus. (a) shows the flower with the segmented lip where the outermost segment imitates the female wasp. (b) A male attempts to copulate with a dummy covered with a synthetic mixture of pyrazines. (c) A male wasp tries to fly away with the false female. This activates the hinge and the wasp is thrown against the column. (d) The process is completed.
Pterostylis banksii from New Zealand is pollinated by solitary bees and bumblebees. The flowers are up to five cm large and have a movable lip (6). 1-3 are sepals, 4-5 petals, and 7 is the ovary. The lip has a hinge at the base and when an insect lands on the lip, its weight will get the lip to swing inwards, and the insect, often a fly, may be held captured for ½-2½ hours before the lip will give in. When the insect is crawling out, it may on its way collect or deliver pollinia. Photo: B Spragg, Flickr.
Anguloa clowesii from Columbia and Venezuela is another orchid with movable lip. The two photos show the lip in its outermost positions.
Many other orchids anaesthetize the pollinator. The South- and Central American genus Coryanthes is probably the most complicated orchid flower, where pollination depends on anaesthetization of a male bee. Two glands at the base of the column drip fluid (red circle) down the bowl-shaped lip. Attracted by nectar scent, the male bee lands on the almost grotesquely large flower (> 20 cm) at the swollen base of the lip. There is no nectar, but the bee then perfumes itself with a hormone-like product produced by the lips basal (‘upper’) part. This makes the male bee attractive to females. However, the scent anaesthetizes the male bee and it falls into the fluid filled lip, where it must fight to avoid drowning. It cannot get a foothold on the lips walls since they are smooth as the inside of a Sarracenia pitcher. Naturally, there is only one way out and that passes the tip of the column with the pollinia and stigma.
The passage past the column is so narrow that it is hard work for the bee to squeeze through but in return it ensures that pollinia are picked up or equivalently adhere to the stigma if the male carries some with it. The stigma is passed first.
As mentioned earlier small flowers are collected in inflorescences in order to make them more visible and social bees pollinate many of them. This is true to e.g. Oncidium truliferum, where the inflorescence is interpreted as a bee swarm. Social bees are also the ones that pollinate many species in the genera Orchis and Dactylorhiza such as D. maculate (left), Orchis morio (middle) and O. mascula (right). The two first mentioned are primarily pollinated by the honeybee Apis mellifera.
Epipactis helleborine is pollinated by the hornet Vespa sylvestris. The nectar partly anaesthetizes the hornet so it has difficulties flying after the visit. Therefore, it tumbles down the inflorescence to the next flower. At right, it is shown falling out of the lowermost flower with pollinia on its head (photo: H Tybjerg). E. helleborine has a long flowering period in July-August and atypically a lasting nectar production. Different wasps, hornets and bees also visit E. palustris.
In Oncidium planilabre (above) and O. hyphaematicum the male bees attack the flower since they interpret the flower as a hostile insect. The bee does not land on the flower but bushes headlong into it in order to chase it away. Photo: Pijl & Dodson 1966.
Listera ovata uses parasitic wasps and bugs as pollinators. The flower has nectar in an open pit on the lip. When the insect hits the rostellum, it explodes and a sticky substance is released which glues the pollinia to the insect. It reacts with fright and leaves the flower immediately but only to sit on another flower, hence the chance of cross-pollination is considerably increased. Flies can also pollinate Listera.
Day-active butterflies pollinate some orchids. In Denmark, this is the case for Anacamptis pyramidalis (left). The flower has a relatively long and very narrow spur, which butterflies can penetrate with the trunk (proboscis). The red flower color is no problem since butterflies can perceive red. The inflorescence is so dense that it as a whole can work as a landing site. The butterfly places itself on the top and turns around itself in order to reach the flowers with the trunk just as shown for Argynnis paphia on the thistle Circium arvense (right). Orchis ustulata (middle) is said to be mostly visited by butterflies. The entrance to the flower is too narrow for bees. On Öland it is seen pollinated by large flies, where the pollinia are deposited on the trunk.
Butterflies of course pollinate the butterfly orchid Psycopsis papilio. There are four species in the genus distributed from the Caribbean to Peru. The flowers emerge successively raised far above the leaves on an up to 70 cm flower shaft where they vibrate in the wind and thereby imitates a flying female butterfly. This attracts male butterflies that try to copulate with the flower and at the same time they perform pollination. The butterfly mimicry of the flower has in addition the effect to attract parasitic insects hunting butterflies where they can lay their eggs.
Platanthera bifolia (left and middle above) is a typical moth pollinated flower. It is white, it smells sweet in the evening, and it has a long spur with plenty of nectar. Since the pollinia are placed closely together at the entrance of the spur, the moth will collect the pollinia on the trunk. In P. chloranthe (middle below) the moth will get the pollinia on the eyes because the pollinia are sitting far apart. Since the stigma in P. bifolia is very small and placed at the spur entrance, it can only collect pollinia placed on the trunk and the two species will only rarely hybridize, although, they are often visited by the same moth. In P. bifolia the stalks of the pollinia furthermore bend forward so the pollinia are simply carried in and out of the spur entrance in P. chlorantha. Also notice the lip. It is turned down and obviously does not function as a landing site, since moths do not land but stand and swirl in front of the flower. – Habenaria dilatata (right) from North America is also moth pollinated.
Aerangis thompsonii with its long spur and the reflexed lip is a typical moth pollinated flower. The same applies to Angraecum eburneum, where the flowers atypically are turned so the lip in this case cannot either function as a landing site, since it is turned upwards. The pollinator stands and swirls in front of the flower and the trunk is guided into the relatively short spur by help of the gutter in the column. Both genera are African.
Angraecum sesquipedale from Madagascar is a noteworthy moth pollinated orchid with an about 30 cm long spur. Darwin described the species in 1862, and from his theories about adaptation and co-evolution between species, he predicted a moth must exist with a trunk (proboscis) of similar length to make pollination possible. Forty years later the right moth was discovered and named Xanthopan morganii f. praedicta. However, it was not until 1997 that it was observed that the orchid was pollinated as predicted. Xanthopan photo: kqedquest, Flickr.
Some orchids are adapted to fly pollination. There are two main types of fly pollination. In the simplest form, the fly has free admittance both to and from the flower. This is the case in e.g. Bulbophyllum the most distributed orchid genus with more than 2000 species. Above B. lemniscatoides is shown, where each tepal is drawn into a long narrow appendix, which doves in the wind and that is attractive to certain flies. Other species smell rotten in different ways, which each attracts a single fly species. – In America the genus Pleurothallus (right) is mainly fly pollinated.
The more advanced type of fly pollination is represented by Paphiopedilum having building features, which resemble other flytraps (pitcher covered by a lid as in Sarracenia, striped lid as in Arisaema, Araceae). Green, brown, dark red, white, and violet are the dominating colors. In addition, some have a bad scent. Flies, beetles, and in some species bees enter through the large opening in the slipper shaped lip and leave the trap through small openings beside each of the two anthers as described earlier for Cypripedium calceolus, which is mostly visited by solitary Andrena bees. Paphiopedilum maudiae var. coloratum is shown at left and the hybrid Paphiopedilum ’Lynleigh Koopowitz’ at right.
Dots and hairiness (indumentum) are also attractive to flies as in Paphiopedilum hirsutissimum (left) from India-Thailand and Cambodia as well as in P. sukhakulii (right) from NE-Thailand.
Rhizanthella gardneri is pollinated by termites. A farmer in SW Australia discovered the orchid in 1928 when suddenly open flowers emerged from the soil while he was plowing his field. The plant grows and flowers completely subterranean. It has no chlorophyll and obtains all nutrients through fungi, which have mycorrhiza with green autotrophic plants. Hence, the orchid is a myco-heterotrophic plant just like species of Monotropa. The about 150 densely packed small flowers attract termites as pollinators and their scent is so strong that even humans are able to sense it through the soil. Only six populations are known, so the species is considered in great danger of extinction. Photo: Jean & Fred. Flickr.
The only group of animals besides insects that play a role in pollination of orchids is birds and American hummingbirds in particular. There is no scent in bird-flowers but strong red, orange, or yellow colors and there are lots of nectar in solid spurs or in nectaries built into the ovary wall. There is no use for a landing site, hence the lip is often narrow or tube shaped as in the Central American Prosthechea vitelline (above) earlier known as Epidendrum vitellinum and Encyclia vitelline (Photo: H Tybjerg). Interestingly, a white form exists in nature, but it is unknown to me if despite its lacking scent it is moth pollinated. Hummingbird photo: K Bondy, Flickr.
Cattleya aurantiaca (left) from Central America with the narrow tube-like orange-yellow lip is reported pollinated by hummingbirds while bees pollinate most other Cattleya flowers. – Sophonitis coccinia (= C. grandiflora, right) from Brazil is also considered pollinated by humming birds.
Cattleya aclandiae growing in the Bahia province in Brazil combines characters for both bird and insect pollination, since the lip functions as a landing site and the flower has a scent while the column forms a red tube suitable for the tongue of a hummingbird.
As mentioned, there are at least 25,000 orchid species besides a number of natural hybrids. Therefore, this review of pollination in orchids describes only a limited part of all the existing variations. It may be added there are at least 20 million of insects of which only 950,000 are described and named. About 9,000 birds are known. If just one of the pollinators becomes extinct, it can easily also cause the disappearance of an orchid species.