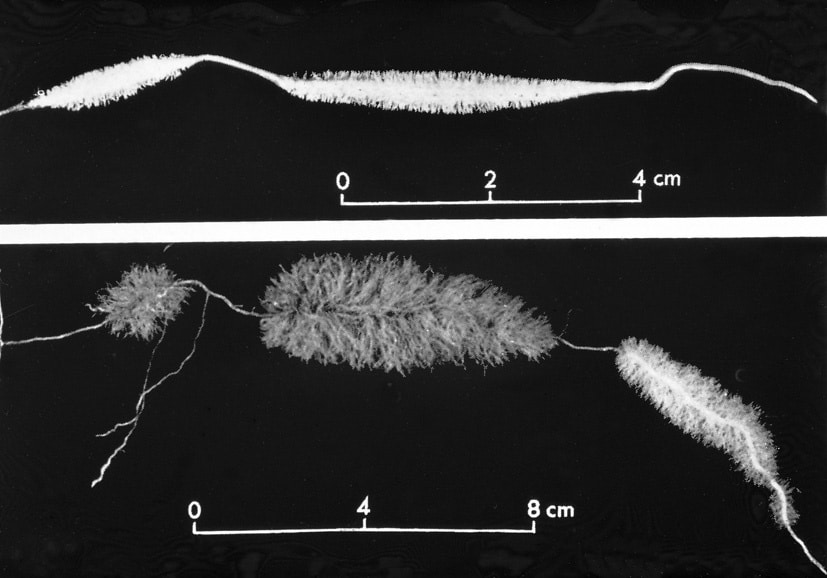
Large absorbing root surface, proteoid roots – shoot/root ratio small.
Microphylli – ericoid and involute or revolute leaves – stem assimilation.
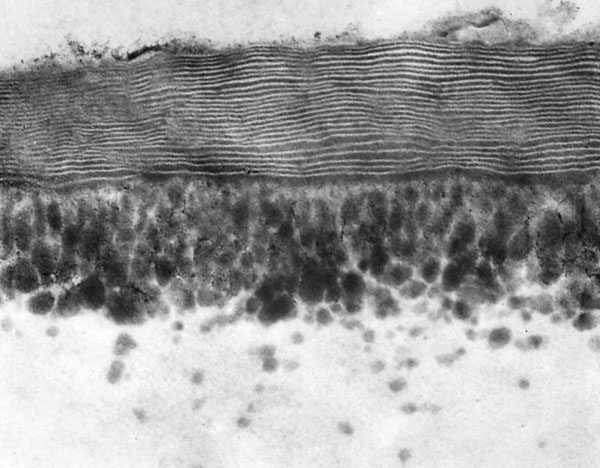
Heavily cutinized epidermis, lamellar cuticle proper reduces cuticular transpiration.
Plenty of epicuticular wax reflects radiation and reduces warming of the leaves.
Indumentum with dead air-filled trichomes increases the boundary layer and reduces warming.
Water absorbing trichomes for absorbing dew and rain.
Ethereal oils in glandular trichomes and cavities reduce surface tension of water and force the liquid phase of water backwards in the cell wall.
Sunken stomata – many stomata/area – permanent closure of superfluous stomata.
Stoma in furrows or cavities reduce transpiration.
Water storage tissue in cortex, marrow, hypoderm, mesophyll (tannin in vacuoles).
Well-developed conductive tissue and mechanical tissue (sclerenchyma), bundle sheath extensions.
Compact assimilation tissue and reduced intercellular airspace minimize area of exposed cell walls.
C4-photynthesis and CAM-photosynthesis.
Some plants endure dehydration better than others, mosses and lichens in particular.
Annual plants cope with the dry season as embryos surrounded be a protective seed coat.