

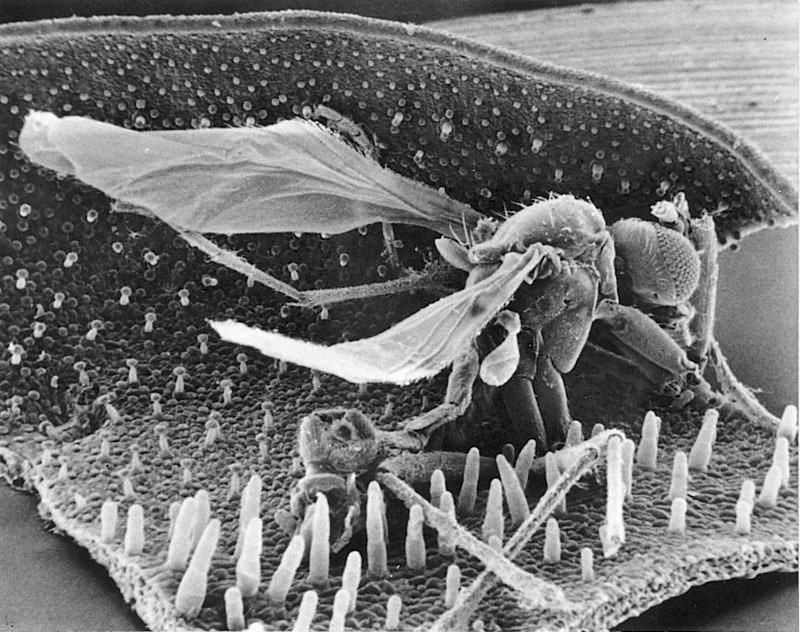
Pinguicula lusitanica with trapped mosquito.
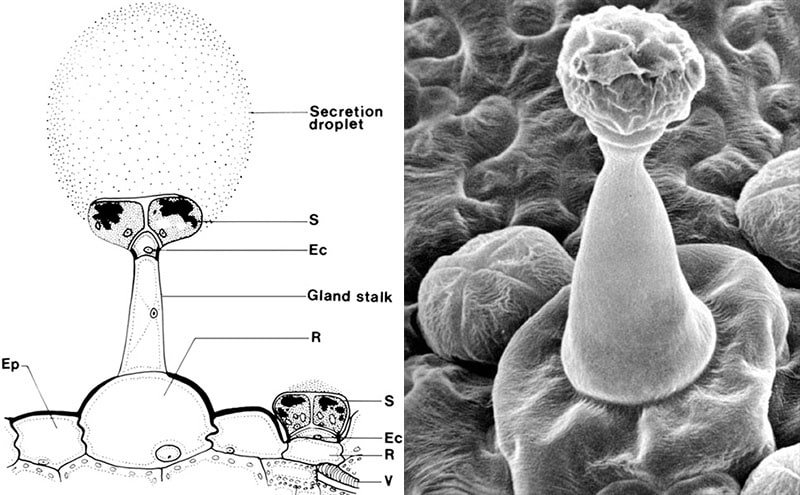
Pinguicula vulgaris. Langstilkede og kortstilkede kirtler. Ec, endodermoid celle. Ep, Epidermiscelle. S, kirtelcelle. R, reservoir-celle. V, Vandledende kar (xylem). Diagram after Y. Heslop-Harrison.

Pinguicula vulgaris with enrolled leaf margins.

Pinguicula moranensis with pool of liquid under the prey.
.
The trap leaves act as a fly paper. The leaves are provided with two types of glandular hairs. Partly long-stalked mucus-secreting glandular hairs built over a ground plan with a basal cell/reservoir cell at the bottom, a stalk consisting of a lower elongated cell and an upper small so-called endodermoid cell and a head with 8-32 gland cells. The endodermoid cell is provided with lipid impregnations in the outer walls, so that all transport in the cell walls is blocked, which gives the cell membrane a controlling function over which substances can be transported through the cell’s cytoplasm. The secreted amount of mucus corresponds to several times the volume of the gland cells. Partly short-stalked submerged glandular hairs built on the same ground plan, but with a very short stalk-cell. The short-stalked ones secrete digestive enzymes and absorb the breakdown products.
It is not known what attracts the insects. Perhaps it is the glistening leaves and/or the faint smell of fungus that often surrounds the plants. In the vast majority of species, a prey of not too large a size will stimulate the leaf to roll the leaf edge towards the prey or to form a small depression under it. These movements are very slow and can take up to two days to complete. The movements are therefore of no importance for the actual catching process, but they cause more of the small seated digestive glands to come into contact with the prey, resulting in faster digestion. The advantage for the butterworth is probably a reduced risk of rainwater washing the prey off the leaves before it is digested.
The digestive glands, which have 2-8 cells in the gland head, are among the smallest of the carnivorous plants. They belong to the dry type, where the secretion of the digestive enzymes only occurs after a chemical stimulation with nitrogenous compounds such as uric acid secreted by the prey. Already one hour after the first stimulation, the secretion of an acidic (pH 3.1-3.4) viscous liquid digestive fluid begins. The liquid also acts as a relaxant, so that the insect’s surface (cuticle) loses its water-repellent properties, and the prey’s respiratory organs are therefore quickly put out of function. After 2-3 hours, the prey is bathed in a pool of secretions, the size of which is roughly proportional to the size of the prey. Each digestive gland can secrete an amount of liquid several times its own volume. Thereby neighboring glands to the first stimulated glands are involved in the secretion pool, and at the same time the stimulation factor is spread, so that the neighboring glands including the long-stalked mucous glands also start a secretion of liquid with digestive enzymes. Presumably, the stimulating substance is eventually used up or diluted, so that the secretion pool has a limited size anyway.
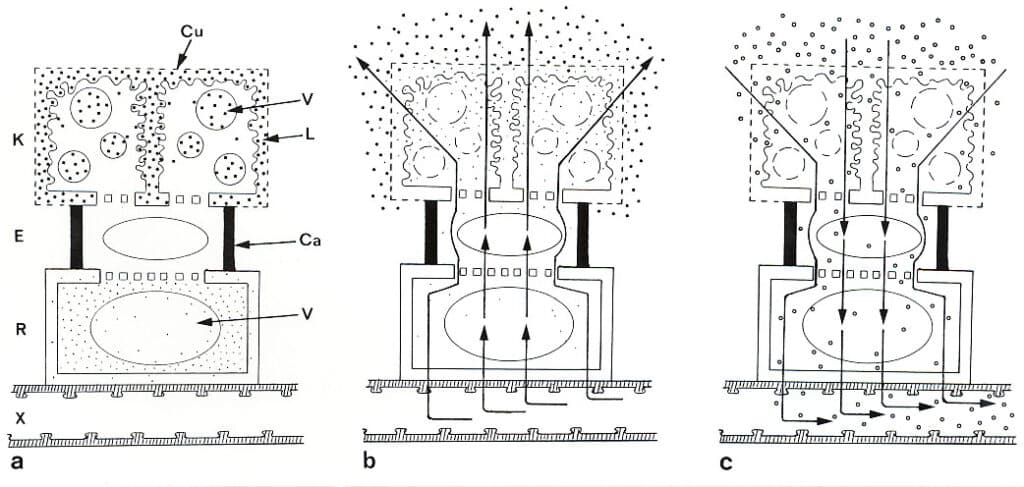
Diagram of functioning digestive gland in Pinguicula (butterworth). a) synthesis/rest phase. b) secretion phase. c) absorption phase. K, glandular cell. E, endodermoid stalk cell. R, basal cell (reservoir cell). X, vessel/tracheid in cord. Ca, lipid blocked (cutinized) cell wall. Cu, perforated cuticle. L, labyrinth wall. V, vacuole. Dots, chlorine ions. Black squares, enzymes. Circles, nutrients. Arrows in b and ce indicate the direction of the water flow. (Based on Y Heslop-Harrison 1975).
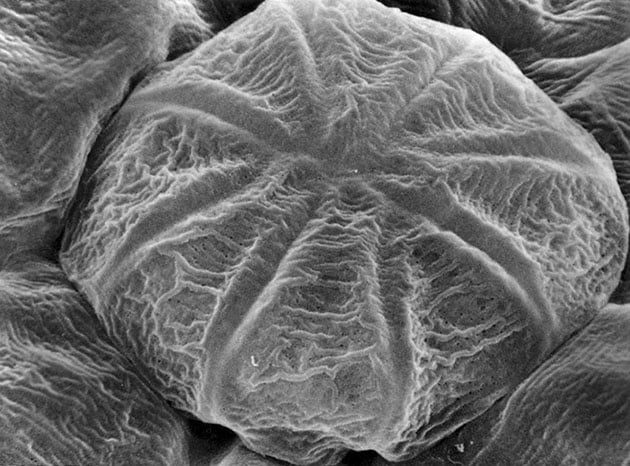
Pinguicula vulgaris. Digestive gland head with eight gland cells. The small dark spots on the surface represent cuticle free areas.
.
.
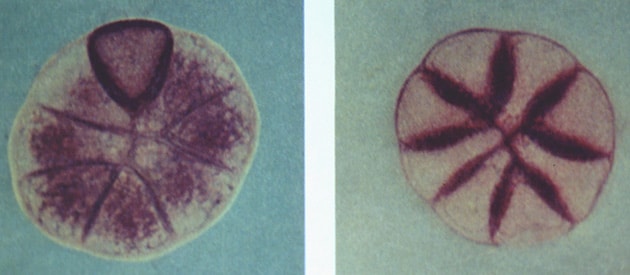
Pinguicula grandiflora. At left, acid phosphatase activity in radial cell walls. – At right, all glandular cells minus one are activated and the enzyme is located primarily in the cytoplasm. Y Heslop-Harrison 1976.
.
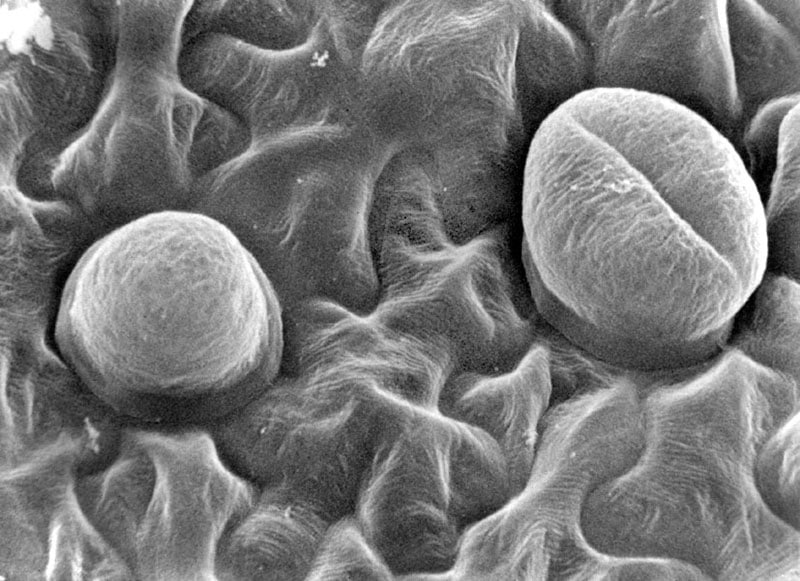
Pinguicula lusitanica, hydathodes on underside (abaxial) of leaf.
|
The digestive enzymes are produced in the cells of the gland head and stored in vacuoles and labyrinth walls. The following enzymes have been found in butterworts: amylase, esterase, acid phosphatase, protease and ribonuclease. At the end of the synthesis phase, a form of controlled degradation (autolysis) of the gland cell membranes takes place, so that vacuoles, cytoplasm and labyrinth walls eventually form one unified enzyme container. Ribonuclease can first be traced in the secretion, and it is therefore thought to be stored in the labyrinth walls. The amylase comes mainly from the long-stalked glands, while the protease comes from the submerged glands. In the resting phase, large amounts of chlorine ions are also stored in the basal cell. The chemical stimulation of the gland cells is supposed to activate an ion pump located in the cell membrane (plasmalemma) of the endodermoid stalk cell, which is well supplied with mitochondria that supply energy for the pump work. The activated pump causes the chlorine ions to quickly flow out into the gland cells, which are now in the secretion phase. The chlorine ions build up an osmotic gradient that will inevitably draw a flow of water out through the gland from the conductive tissue. This water flow flushes the stored enzymes out of the gland, as the cuticle of the gland cells is perforated, as in sundew (Drosera), so that both ions and molecules can pass in and out unimpeded. In certain places in Sweden, the insides of thick milk bowls have been smeared with the slimy leaves of Pinguicula vulgaris, after which the protein-splitting enzymes promoted the precipitation of cheese substances.
In the absorption phase, the water flow reverses. This happens partly because the powerful pumping action destroys the plasmalemma in the endodermoid stalk cell, so that it loses the ability to control the direction of the current, partly because during digestion a high concentration of nutrients builds up in the secretion pool. In relation to this, both the ion concentration and the water pressure in the tracheids and vessels of the conductive tissue will be so low that the water flow is forced to reverse according to the laws of physics on diffusion. On the underside of the leaves there are some water-secreting glands (hydathodes) which may also play a role in controlling the flow of water to and from the digestive glands. As the majority of membranes gradually break down, the digestive glands can only be used once, but this is compensated for in e.g. P. vulgaris which during the growth period produces a new leaf every fifth days. It will be approx. 400 cm2 adhesive paper per growing season.