









Skadelige snylteplanter
Fra en botanisk synsvinkel er snylteplanter også kaldet parasitiske blomsterplanter fantastiske planter. Mange viser stor skønhed i blomsterne, hvor sofistikerede mekanismer til bestøvning og spredning kombineres i Loranthaceae, mens ekstremt reducerede og usædvanlige vegetative og florale strukturer forekommer i holoparasitiske familier som Hydnoraceae, Rafflesiaceae og Balanophoraceae. Både fra et æstetisk og et biologisk synspunkt må man beundre disse planter, der har specialiseret sig i at udnytte værtsplanter som delvis eller fuldstændig næringskilde. Samtidig har nogle snylteplanter dog en betydelig negativ indflydelse i varmere klimaer på millioner af menneskers liv og økonomi, og den primære opgave for flere forskningsinstitutioner er at studere og udvikle metoder til bekæmpelse af disse skadelige snyltere. Denne side omhandler de mest skadelige parasitter samt, hvad der gøres for at bekæmpe dem. Se siden Snylteplanter for en oversigt over samtlige familier. – Billeder og diagrammer uden navn på leverandøren er produceret af forfatteren. Titelfoto: Arceuthobium douglasii med hanblomster.
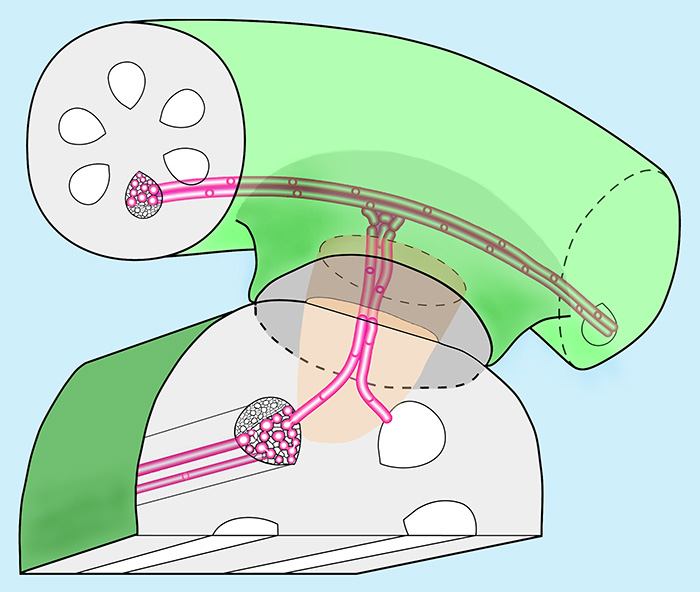
En snylteplante står i fysisk forbindelse med én eller flere værtsplanter via ét eller flere haustorier. T.v. vises et diagram over et fungerende haustorium. Parasitten er øverst og værten nederst. Parasitten anlægger en hæfteskive, der limes til værten. I hæfteskiven dannes et vækstpunkt, der producerer et intrusivt organ (sænker – lysebrun), der ved enzymatisk opløsning af cellevægge og mekanisk tryk vokser ind til værtens ledningsstrenge, hvorefter de to parters xylem celler (vandledende karceller og tracheider) forbindes til en xylembro (rød på figuren). Hos de mest avancerede er kontaktfladen åben som på transmissionsmikroskop billedet (Cassytha, efter Damachke 1982). På farvebilledet er den primære sænker hos mistelten (Viscum album) ramt perifert. Øverst ses flere haustorier (snylter øverst) anlagt af Triphysaria versicolor fra gyvelkvæler familien (Orobanchaceae).


Haustorier kan variere meget i både form, størrelse og antal. T.v. ses et meget stort primært hausorium udviklet direkte fra kimroden hos Loranthecéen Erianthemum ngamicum fra Sydafrika. Bemærk hvor tynd værtsgrenen er på ydersiden af haustoriet som et tegn på, hvor effektivt haustoriet har drænet værten for vand og næring. T.h. ses både primært haustorium (P) og sekundære haustorier (S) anlagt fra en epicortikal rod (adventiv rod fra stænglen) hos Notanthera heteropylla (Foto: J. Kuijt).
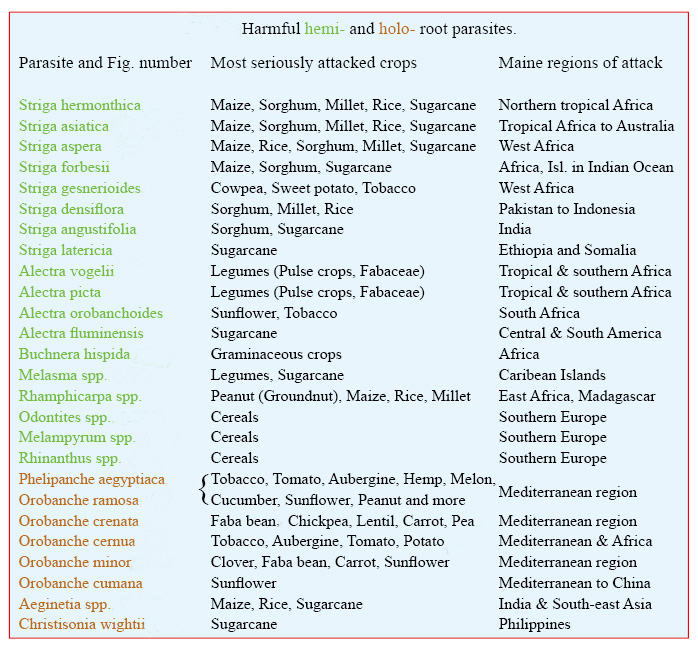
Af de omkring 4.500 arter af parasitiske blomsterplanter er det kun meget få, der parasiterer landbrugsplanter. Slægterne Cuscuta, Striga og Orobanche er de vigtigste. Ved angreb i frugtplantager og skovbrug kan stængelparasitter fra Loranthaceae og Viscaceae tilføjes, især arter af Arceuthobium. Tabellen er på engelsk, da den er taget fra min bog ’Parasitic Flowering Plants’. Tabellen angiver de værste rodparasitter, hvilke afgrøder de angriber, og i hvilke områder angreb har størst skadelig effekt. For hver slægt nævnes først den art, der forårsager flest problemer og størst økonomisk tab. Tre vigtige lærdomme fremgår af tabellen. For det første angriber langt de fleste skadelige parasitter mere end én afgrødeart. For det andet kan næsten alle afgrødearter angribes af mere end én parasitart. For det tredje undgår Latinamerika næsten skadelige parasitplanter.

Før vi ser på omfanget af de skader, parasitterne kan forårsage på dyrkede planter, er det relevant at se på, hvordan vilde værter reagerer. Snylterens træk på vand og næringsstoffer fra værten vil ofte reducere værtens livskraft. Derfor hæmmes den vegetative vækst, og værten producerer færre blomster, frugter og frø. En parasit er dog sjældent stærk nok til at dræbe værten. Striga dræber værten på én vækstsæson, men for andre vil det tage flere eller mange år. Det gælder også for ikke-naturlige værter, som f.eks. når Viscum minimum dyrkes på Euphorbia obesa (herover). Den sunde vært t.v. er fotograferet 17 mdr. efter såning af V. minimum, og nogle få små grønne skud ses. Planten t.h. er fotograferet godt 3 år efter såning af V. minimum, og snylteren er brudt frem talrige steder, mens værtens ribber tydeligvis er sunket sammen, og værten er kraftigt svækket. At dræbe værten er helt klart en ulempe for parasitten. Énårige parasitter kan dog have råd til denne luksus, fordi de overlever til den næste sæson som frø. Her er der således en vigtig grund til, at énårige rodparasitter, der producerer talrige frø, er så succesfulde og skadelige for dyrkede planter.


Det gælder generelt, at hvis værten har det godt, vokser dens parasitter sig også stærkere. Sammenlign størrelsen og antallet af blomster på den fuldt udviklede Orobanche elatior, der snylter på stor knopurt (Centaurea scabiosa), der vokser i næringsfattig kalksten (Fig. t.v. hvor en beskeden værtsplante ses uskarpt midt i billedet), med den samme art, der vokser i mere næringsrig jord (Fig. t.h. Foto: H. Madsen). Det er tydeligt, at en velnæret vært er en fordel for en parasit, som derefter har tendens til at være mere aggressiv. Afgrøder er generelt velnærede, og konkurrerende ukrudt fjernes ofte enten mekanisk eller med pesticider. Nogle rodparasitter, der i naturen lever af græsser såsom Striga eller foretrækker bælgfrugter såsom gyvelkvæler (Orobanche), har i afgrøder fundet velnærede værter i stort tal, der tillader parasitterne at udvikle sig ekstremt godt, sætte masser af frø, og derfor bliver disse parasitter virkelige skadevoldere.
Snylteplanter i landbrug


Udover Cuscuta og Cassytha er det rodparasitter fra Orobanchaceae, der forårsager problemer i landbruget. Lejlighedsvis angriber rodparasitter fra Santalaceae også afgrøder. I Sydeuropa kan en art nålebæger Thesium humile (Fig. t.v. Foto: José Quiles) angribe hvede og byg, mens andre Thesium arter kan angribe sukkerrør i Australien og Afrika. Figuren t.h. viser durra (Sorghum) angrebet af rødblomstret Striga asiatica og pinkblomstret S. hermonthica (Foto: A. Larsen). Dette dobbelte angreb øger kun vanskelighederne med at dyrke f.eks. majs, durra, ris, sukkerrør eller bælgfrugter inden for udbredelsesområdet af skadelige Striga– og Orobanche arter.
Cassytha og Cuscuta (nældesilke)

Cassytha er den eneste parasitiske slægt i Laura-ceae, der ellers består af autotrofe planter. De fleste forskere accepterer denne systematiske holdning, men andre har foreslået en separat monotypisk familie, Cassythaceae. Det er en lille slægt med 17 arter, hvoraf 14 findes i Australien, mens én art, C. filiformis, er pantropisk. Cassytha vokser overvejende kystnært. Især C. filiformis er ofte en del af strand vegetationen (Fig. t.v. Foto: P. Vestergaard), men findes også hundrede km eller mere inde i landet. Enhver plante kan bruges som vært.


Cassytha arter er flerårige og praktisk talt bladløse slyngplanter med stængler, der drejer til venstre dvs. mod uret (Fig. t.v.) omkring alle dele af de støttende værtsplanter. Det samme er tilfældet hos nældesilke Cuscuta. Nogle engelske lærebøger kalder dog stænglerne i begge slægter for højresnoede. Der er et skifte mellem tætte vindinger (Fig. t.v.) med sekundære haustorier og lange haustorieløse stængler, der rækker ud efter nye værtsgrene. Derfor ses stænglerne ofte hængende frit i luften som på Fig. t.h. af Cassytha filiformis. Billedet illustrerer samtidig, hvor kompakt Cassytha kan optræde og derved skygge værtsgrene ihjel. De sekundære haustorier kan opfattes som modificerede adventivrødder, men er måske snarere en unik struktur (sui generis), da der ikke kendes adventivrødder hos andre arter i Lauraceae. De slyngende stængler indeholder slimkanaler, der måske gør stænglerne mere fleksible.


T.v. er en hæfteskive fra Cassytha pubescens gjort fri af værten, og et kileformet intrusivt organ er brudt frem. T.h. sidder hæfteskiven på værten, og det intrusive organ er vokset frem fra hæfteskivens indre, hvor flere cellelag er presset sammen under organets vækst.


Cassytha arter skaber kun problemer lokalt i tropiske og subtropiske områder og hovedsageligt på træer. Den mest udbredte art C. filiformis (herover) kan angribe mango, citrus, kaffe, kakao, te, kokosnød, papaya, og forskellige prydplanter. Skyggeeffekt og vandoptagelse fra værten resulterer i tab af blade og reduceret fotosyntese, og det giver mindre udbytte. De største problemer forårsager arten i Vestafrika og Caribien, hvor den angriber kokos og unge mango- og citrusplanter og hurtigt spreder sig over store arealer. Blomsterne er små og uanselige, mens frugten er et énfrøet bær, der hos nogle arter er spiselig og hos andre stærkt giftig. Hos nogle arter er frugtskallens inderste lag (endocarpiet) dog så hårdt, at frugten bliver en stenfrugt.

I Australien kan Cassytha pubescens (t.v. Foto: R. Hotchkiss) være et problem på marker med vindruer, hindbær, blåbær og prydplanter. Det kan føre til nedsat vækst og dårligere frugt kvalitet. Det giver økonomiske tab især på vinmarker, hvor omkostninger til manuel fjernelse og bekæmpelse med herbicider desuden stiger.


Vegetativt ligner Cassytha og nældesilke Cuscuta i overraskende grad hinanden. De er begge halvparasitiske, bladløse, stængelassimilerende, venstresnoede slyngplanter. De anlægger kun sekundære haustorier, og haustoriernes anlæggelse er meget ens, men den florale struktur er meget forskellig med 6 blosterblade hos Cassytha (t.v. Foto: C. Gracie), mens Cuscuta er 5-tallig (t.h.). Det viser, de hører til to forskellige familier Lauraceae og Convolvulaceae/Cuscutaceae, hvor begge er eneste parasitiske slægt i familien. De to slægter er således et klassisk eksempel på konvergent evolution. Der er dog forskelle, mens Cassytha er flerårige er de fleste Cuscuta énårige, og deres fotosynteseevne er ofte reduceret, og enkelte arter som vores hjemlige nældesilke (Cuscuta europaea) er endda holoparasit uden fotosyntese. Cuscuta betragtes derfor som mere avancerede parasitter.



T.v. ses kraftig blomstring hos Cuscuta europaea efter angreb på Rejnfan (Tanacetum vulgare). Arternes hurtige vækst i begge slægter kan tilskrives det store antal haustorier, hvor begge anlægger en xylembro med åben kontakt til værtens ledningsvæv (se Cassytha øverst på siden). Hos Cuscuta sker der yderligere det, at xylembroens kontaktceller (som vist herover) invaderer værtens vandledende tracheider (kendes på de spiralformede afstivningslister på cellevæggens inderside).


Cuscuta haustorier er yderligere avancerede ved, at de sammen med arter fra Orobanche er de eneste, der foruden en xylembro også har kontakt til værtens sivæv (phloem). Det er vist skematisk på figuren t.v., hvor en parasitcelle med fingerformede forlængelser griber omkring en værts sirørscelle, der bl.a. transporterer fotosynteseprodukter. Den røde indramning og det elektronmikroskopiske foto t.h. viser samme situation (S, sirør. P, parasitcelle). Snylterens kontaktcelle har udviklet en labyrintvæg på kontaktfladen, der forøger cellemembranens areal betragteligt, og det indikerer intensiveret overførsel af næringsioner fra vært til parasit, der er medvirkende til, at de énårige planter kan vokse så hurtigt. Diagram og TEM foto, I. Dörr.


Billederne herover fra Baha California illustrerer, hvor hurtigt Cuscuta arter kan vokse. T.v. er det C. salina på en kveller Salicornia sp. og t.h. er det C. veatchii på Bursera sp. Det havde ikke regnet i 6 år i området, men 5 uger inden et besøg i 1990 regnede det, hvorefter de viste snylteplanter spirede og udviklede sig til store tætte måtter. Man har beregnet, at en Cuscuta plante fra et enkelt frø kan producere mere end 1,2 km stængler. Billedet t.h. indikerer, at fugle har været involveret i frøspredning samt lang tids bevaring af spiringsevne. Det kan forklare tilstedeværelsen af C. veatchii i toppen af 8 m høje værtstræer, da det er usandsynligt, at snylteren kan nå trætoppene på under 5 uger ved spiring på jorden.


Cuscuta arter angriber primært urteagtige afgrøder i både tempererede og tropiske områder. Omkring 20 arter af Cuscuta er registreret fra afgrøder, og de arter, der oftest forårsager problemer, er den nordamerikanske C. campestris (Fig. t.v. Foto: D. M. Joel), som er blevet introduceret i mange lande verden over gennem frøkontaminering. Den ses her gøre skade på tomatplanter. T.h. har C. epithymum angrebet lucerne (Foto: B. O. Lyshede). Af andre arter, der ofte kan volde problemer på markafgrøder kan nævnes C. australis, C. chinensis, C. reflexa, C. monogyna, C. lupuliformis, C. japonica, C. pedicellata, C. planifolia, C. gronovii og C. indecora.
Blandt angrebne afgrøder foruden tomat og lucerne er nigerfrø (Guizoltia abyssinica) og gulerod på grund af samme frøstørrelse, der gør det vanskeligt at rense frøene. Cuscuta forårsager også problemer for avlere af aubergine, løg, sukkerroer, sojabønner, kartofler, krysantemum og tranebær. I Californien reducerer 20% angrebne tomat planter udbyttet med 25% og forårsager et tab på 4 millioner dollars/år. Generelt har Cuscuta lav værtsspecificitet, dvs. de accepterer mange værtsarter, men C. epilinum har forholdsvis høj værtsspecificitet og er hovedsageligt kendt fra hør, men også fra lucerne, som ellers er mest truet af C. campestris.
Udbyttetab synes at være mest alvorligt i Østeuropa og asiatiske lande hos lucerne, kløver og sukkerroer og i Nordamerika hos lucerne. Det anslås også, at sandsynligvis mere end 100.000 ha sojabønner (Glycine max) er angrebet i Kina. I Indien forårsagede Cuscuta campestris et udbyttetab på 85,9% i nigerfrø, 81,6% i grønkålsbønner (Phaseolus radiatus), 66,8% i sesam (Sesamum indicum) og 48% i sojabønner, men intet tab i ris (Oryza sativa), fordi sclerenchym (styrkevævceller med tykke forveddede vægge) i stænglerne forhindrer haustorier i at etablere sig. I potteforsøg med Cuscuta chinensis på tomat blev frugtudbyttet reduceret med 72%. Skøn over udbyttetab i sukkerroer inficeret med C. campestris er i størrelsesordenen 3500-4000 kg/hektar. I lokale områder er et totalt udbyttetab muligt, og angrebne foderplanter må til tider opgives, fordi Cuscuta kan være mildt giftig for husdyr.
Der er flere metoder til bekæmpelse, men de fleste er ret ineffektive eller besværlige. Der arbejdes på biologisk bekæmpelse med svampe og bakterier, men stort set kun på forsøgsstadiet. Rensning af frø hos især tomat, lucerne og gulerod samt brug af certificeret frø er nyttigt ligesom manuel fjernelse af snylteren og ukrudt, der kan fungere som vært og rengøring af redskaber, der kan medbringe frø. Der følger senere en mere detaljeret gennemgang af bekæmpelses metoder.
Striga

Striga hører til gyvelkvæler familien (Orobanchaceae), og kortet viser slægtens udbredelse. I området inden for de røde linjer forårsager Striga arter de mest alvorlige skader på dyrkede planter. De to røde pletter i USA markerer, at S. asiatica er introduceret i Carolina og S. gesnerioides er introduceret i Florida.


Striga kaldes på engelsk witch weed, sandsynligvis fordi flere arter på trods af deres skønhed synes at udføre ’ond magi’ som en heks. Heksene i den antikke verden blev kaldt striges, og en striga var en bloddrikkende natteånd, der sugede blod af børn, mens Striga suger saften af deres værter. De omkring 40 arter er hjemmehørende i Afrika, Asien og Australien. Striga arter er etårige eller sjældent flerårige, altid med et primært haustorium og hos nogle arter også med sekundære haustorier. Bladene er næsten modsatte, og nogle som S. hermonthica har prangende blomster (Fig. t.v.). Bestøvning sker af insekter, sandsynligvis mest sommerfugle. På Fig. t.h. besøger en Eurema sp. S. asiatica (Foto: J. Visser). Selvbestøvning er dog blevet observeret i en stamme af S. asiatica introduceret i USA, hvor pollen falder ud af støvknapperne ned på støvfanget, før blomsten åbner sig. Frøene er små omkring 0,3 mm, produceres i stort antal og spredes hovedsageligt af vinden, men de klæber også sammen med mudder til klovdyr, hovdyr, redskaber og fodtøj og spredes derved på dyrkede marker.


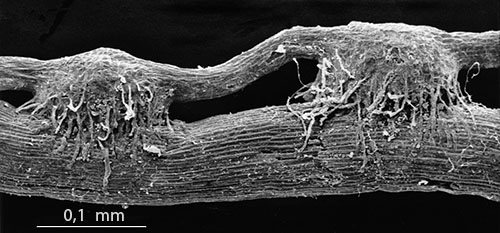
Efter frøspiring stimuleret af udskillelser fra værten danner kimroden enten først en hæfteskive som hos Striga hermon-thica (Fig. t.v., Foto: I Dörr) eller vokser direkte ind i værten. Nogle arter anlægger desuden en knold (tubercle) fra kimroden, og herfra skyder de blomstrende skud frem. De xylemelementer, der er i kontakt med værtsxylemet, er af kartypen, og hos S. hermonthica, og sandsynligvis også hos andre arter, vokser de gennem porefelter i karcellerne og danner åbne skorstene (osculae) inde i værtsxylemet (Fig. t.h., Foto: I. Dörr). Transportmodstanden er derved næsten fuldstændigt elimineret på grænseflade niveau og er med til at forklare snylterens hurtige vækst.

Angreb af Striga er et alvorligt problem i den gamle verdens tropiske og subtropiske regioner (se kort ovenfor). Skal en enkelt blomstrende plante udnævnes til den mest skadelige ukrudtsplante i verden, må det være rodparasitten Striga hermonthica (Fig. herover viser visne og døde majs planter). De næste 5 eller 6 Striga arter i tabellen (klik højre foto) forårsager også alvorlige problemer i flere lande, og 11 arter er kendt for at angribe afgrøder. FN-organisationen FAO anslog i 2008, at S. hermonthica i de seneste år forårsagede et udbyttetab i Afrika på mindst 40%, hvilket repræsenterer over 7 milliarder US dollars årligt. Lokalt kan udbyttetabet være 100% (Fig. t.h). Dertil kommer tabet fra mange af de andre skadelige parasitter anført i tabellen, hvilket betyder, at to tredjedele af Afrikas 73 millioner hektar afgrøder er truet af parasitiske planter, og over 100 millioner menneskers liv i Afrika er alvorligt påvirket i 17 lande og moderat påvirket i yderligere 8 lande. Tal for 2024 viser, at bekæmpelse ikke har hjulpet meget, da tab på op til ca. 80 % stadig forekommer i produktion af perlehirse, og at tab på 30 til 100 % ved dyrkning af majs og durra (Sorghum) forekommer syd for Sahara og i Østafrika.
En plante af Striga hermonthica kan producerer 100.000-500.000 små frø, som spredes på flere måder. De fleste frø spredes sandsynligvis af vinden, men oversvømmelser, kvæg, mennesker og landbrugsredskaber er andre faktorer. Våd jord eller mudder med frø vil klæbe til klove, hove, fødder, sko og redskaber og blive transporteret rundt fra mark til mark. Regnvand vil dræne frøene ned i den øverste jord, hvor spiring finder sted efter påvirkning fra stimulerende stoffer produceret af værtens rødder. Forskellige lokale sorter af majs, durra og hirse dyrkes i forskellige lande, og nogle sorter viser mere resistens over for parasitten end andre. Dette kan forklares med variationer i typen og mængden af spirings stimulerende stoffer, der findes i rodexudater fra de forskellige afgrødesorter, eller med forskelle i andre resistensmekanismer såsom vævs-inkompatibilitet. Så snart det primære haustorium er etableret, sker udviklingen hurtigt. Når blomstringsskuddet observeres over jorden, er der allerede trukket så mange næringsstoffer ud af værten, at det er for sent at gøre noget for at undgå udbyttetab. Der kan dog gøres noget, før haustoriet er etableret, som beskrevet senere under Bekæmpelsesmetoder.

Kemisk struktur af et par kendte spiringsstimulanter. Strigol fra
bomuld (Gossypium hirsutum) og SXSg fra durra (Sorghum). Kilde ?
Når et parasitfrø spirer, reagerer kimroden på de kemiske spirings stimulerende stoffer ved at vokse direkte mod værtsroden for at opnå kontakt. Dette blev først observeret hos Orobanche i 1823. De spiringsstimulerende stoffer bryder frøets dvale tilstand. Forskningen har mest koncentreret sig om stimulanter, der er effektive mod de skadelige Striga arter. Den kemiske struktur af de to mest kendte er vist i diagrammet. (+)-Strigol er et rodexudat fra bomuld (Gossypium hirsutum). Det er effektivt på flere slægter såsom Striga, Alectra og Orobanche på trods af, at bomuld ikke er vært for nogen af disse parasitter. Det strukturelt meget forskellige SXSg (Sorghum Xenognosin for Striga spiring) er et rodexudat fra Sorghum, der siges at være effektivt mod Striga asiatica, som naturligt angriber Sorghum arter. Rodexudaterne er kun effektive inden for kort afstand fra værten, da de hurtigt fortyndes af vand i jorden, og kemiske ændringer sker også ved oxidation. Den kritiske afstand eller effektive zone for stimulanterne er kort, og parasitfrøene skal lande inden for få millimeter fra den potentielle værtsrod eller være til stede i jorden og vente på, at en rod kommer tæt nok på. Eksperimenter med spiring af frø af Striga asiatica i agar har vist, at den kritiske afstand i denne situation er 3-5 mm. I naturlig jord er den kortere. Strategien kræver også et stort antal små, langlivede frø for at øge sandsynligheden for, at nogle frø lander inden for den kritiske afstand. En enkelt S. asiatica plante kan producere omkring 50.000–100.000 frø, og hvert frø er 0,1 mm stort og vejer kun ca. 2–6 µg. Spiringsevnen bevares i mere end 10 år.



Striga asiatica er en meget variabel art med både rød- og gulblomstrede former. Foto: D. C. H. Plowes.
Striga gesnerioides. Foto: A. Larsen.
Striga asiatica findes både i Afrika og Asien. Den har græsser og star arter som naturlige værter, og det er derfor ikke overraskende, at den angriber dyrkede planter som majs, durra, ris, hirse og sukkerrør. De fleste græsværter har C4-foto-syntese (forklaret på siden Xerofytter), ligesom det også er tilfældet med de nævnte afgrøder, undtagen ris. Arter, der forekommer i det køligere højland, såsom S. asiatica, har græsser med C3-fotosyntese som vært over 2.000 m.o.h. S. asiatica er registreret op til 2.480 m. På savannen vil et enkelt værtsindivid normalt blive parasiteret af blot en enkelt parasit. Dette angrebsniveau er ikke et alvorligt problem for værten, da den vil overleve. På dyrkede marker er værtsplanter til stede i stort antal, og deres vækstbetingelser er normalt optimerede. Desuden har den stigende menneskelige befolkning gjort det nødvendigt at intensivere arealanvendelsen, monokulturer er blevet mere almindelige, og sædskifte med ikke-værtsplanter samt lange perioder med braklægning er reduceret. Resultatet har været mere kraftige Striga planter, øget frøproduktion og mange flere end én parasit pr. værtsplante. Denne onde spiral har tvunget mange afrikanske landmænd til at opgive deres gårde i slutningen af det 20. århundrede.
Striga gesnerioides, der er udbredt i Vestafrika, har også græsser og star arter som værter, men går desuden på ærteblomstrede planter, og det går ud over afgrøder som vignabønner (Vigna unguiculata, hvoraf der er fire dyrkede underarter), søde kartofler, men også tobak. Det er især et problem for vigna bønner i Mali, Burkina Faso, Niger, Nigeria, Tchad og Cameroun. En anslået gennemsnitlig reduktion i udbytte på 30% synes realistisk, men på mange marker opleves et totalt afgrødetab. Stærkt angrebne værtsplanter har et reduceret bladareal og udvikler nekrotiske pletter på bladene, som derefter har reduceret fotosyntese og også visner tidligt og falder af. S. gesnerioides forekommer også i det sydlige Afrika, men i en anden sort eller stamme, der ikke angriber vigna bønner, men kun vilde bælgfrugter. Siderødder udvikles ikke, og der er derfor ingen sekundære haustorier. For de øvrige skadevoldende Striga arters forekomst henvises til tabellen, men det kan nævnes, at hele seks af de skadelige arter forekommer i Etiopien, Striga hermonthica, S. asiatica, S. latericia, S. aspera, S. gesnerioides og S. forbesii.
Andre halvparasitiske Orobanchacéer



Alectra er en slægt med 40 arter med primær udbredelse i tropisk og subtropisk Afrika, dog findes A. fluminensis i Mellem- og Sydamerika. Der er både én- og flerårige arter. Det er som de øvrige Orobanchacéer rodparasitter, og specielt A. vogelii (herover på Vignabønne. Foto: M. Hyde) og A. picta volder problemer på bælgplanter i det sydlige Afrika. Udbytte-tabet ved produktion af vignabønner (Vigna unguiculata) varierer fra 20% til 100% og kan sammenlignes med tabene fra Striga gesnerioides, som i det sydlige Afrika erstattes af Alectra vogelii. Landmændene forlader deres marker, hvis der opstår et alvorligt angreb, og det kan ske fra år til år, da en enkelt plante producerer fra 400.000 til 600.000 frø. Alle slags bælgplanter undtagen dueært/ærtebønne (Cajanus cajan) er truet. Vignabønner inficeret med A. vogelii har en tendens til at visne, selv før parasitten viser sig over jorden. Angrebet resulterer i forsinket og reduceret blomstring og dermed reduceret frøsætning. Den tidlige visnen forekommer selv hos godt vandede planter. Der er dog også lidt positivt ved A. vogelii, for i det sydlige Afrika har lokale i mange år brugt de nederste stængeldele til at få et lysegult naturligt farvestof, der kan påføres tøj og flettede kurve.



Der er ca. 100 Buchnera arter, og her vises t.v. B. longispicata fra Zimbabwe (Foto: B. Wursten) og i midten B. americana fra Florida (Foto: C. Gracie). Buchnera arter er relativt harmløse, men B. hispida angriber majs og durra i Nigeria. T.h. ses Melasma scabrum fra Zimbabwe (Foto: D. C. H. PLowes) med 3,5 cm lange blomster. Der er syv flerårige arter fordelt på det sydlige Afrika og tropisk Amerika. De synes kun at være et problem i Caribien, hvor flere arter angriber bælgplanter og sukkerrør.


Rhamphicarpa er énårige hemiparasitiske rodparasitter med små hvide blomster. Der er mindst 6 arter, hvoraf den mest skadevoldende R. fistulosa (t.v.) er udbredt i tropisk Vestafrika og Østafrika inklusive Madagaskar, og den findes også i Tyrkiet, Indien og på New Guinea og N-Australien. Blomsterne åbner sig først om aftenen og bestøves formentlig af natsværmere. Den foretrækker vandmættet jord og kan gøre betragtelig skade i rismarker, men angriber også majs, hirse og jordnød. Den på frimærket viste R. montana’s systematiske placering er usikker, og den anbringes af mange hos Cycnium.



De tre viste arter herover Mark rødtop (Odontites vernus = O. verna), Liden skjaller (Rhinanthus minor) og Ager kohvede (Melampyrum arvense) er hemiparasitiske rodsnyltere, der alle forekommer i Danmark, men det er kun i Sydeuropa, de lokalt volder skade på kulturplanter, især grøntsager på mindre frugtbar jord. Alle udvikler sekundære haustorier på mange værtsarter. Græsser er de vigtigste værter hos alle tre slægter. Der er 30 arter i slægten Odontites. De fleste er énårige, men enkelte er dværgbuske. Der er 45 énårige Rhinanthus arter. De fleste vokser på enge, bestøves af humlebier og har vindspredning. Et par arter er introduceret i USA. Der kendes 35 énårige Melampyrum arter. Frøene har form og størrelse som hvedekorn, deraf det danske navn kohvede. Planterne spises af kvæg, men frøene er giftige for mennesker. Frøene har et olieholdigt vedhæng (elaiosom) som tilpasning til myrespredning.
Orobanche - gyvelkvælere

O. crenata
O. flava
O. californica
O. uniflora
Gyvelkvælerfamilien Orobanchaceae er udbredt over hele verden. Familien består med en enkelt undtagelse af parasitiske slægter. Undtagelsen er den autotrofe Lindenbergia, der betragtes som den mest basale slægt i familien, fordi den repræsenterer den oprindelige, ikke-parasitære tilstand. Molekylære studier har medført, at alle parasitiske slægter, der tidligere var placeret i maskeblomstfamilien (Scrophulariaceae), således også Striga, nu er inkluderet i Orobanchaceae, der rummer 88 hemiparasitiske slægter med ca. 1800 arter og 17 holoparasitiske slægter med ca. 270 arter. Om de ’fredelige’ danske og nordiske arter se her.


Orobanche arter er alle holoparasitiske rodparasitter med en avanceret haustorie bygning. Der anlægges som nævnt for Striga en knold (tubercle), hvorfra blomsterstanden udvikles, når der er oplagret nok næring. På billedet t.v. af O. uniflora skyder en enkelt blomsterstand frem fra tuberclen og efterlader et ar, hvor knoldvævet er sprængt (rød pil). Alle Orobanche har et primært haustorium markeret med rød pil på figur t.h. af O. hederae, der har udviklet tre blomsterstande fra tuberclen. Flere arter danner små sekundære haustorier fra de meget korte rødder, der udgår fra tuberclen. Det primære haustorium er meget effektivt til at dræne værtsroden for vand og næring, og på billedet ses, at værtroden er indskrumpet og død efter haustoriet.

{kind=link}
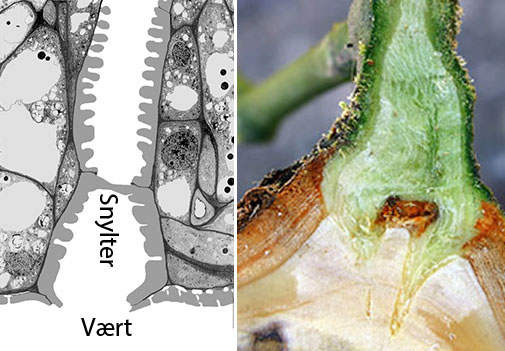
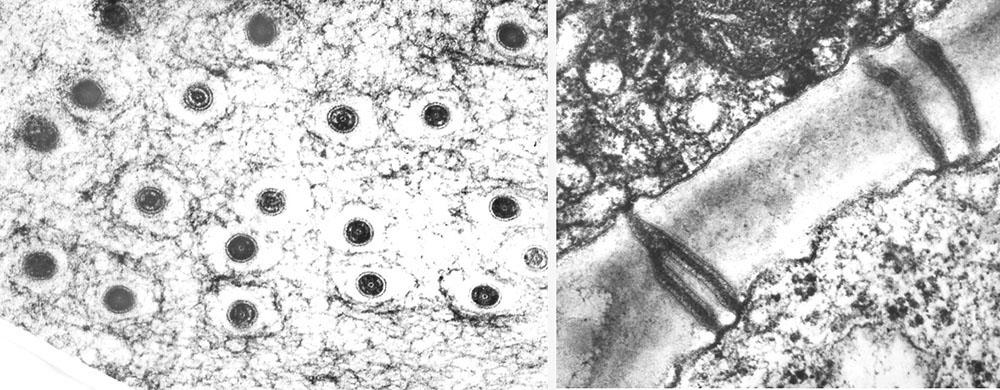
Planten herover, Orobanche uniflora (=Aphyllon uniflorum), fra Vancouver Island snylter på stenbræk (Saxifraga) og sten-urt (Sedum) og skader ikke kulturplanter, men haustoriet er bygget som hos de skadelige arter. T.h. er dele af det primære haustorium set i et tværsnit af værten. Grænsefladen (interface) mellem snylter og vært er markeret med en gul linje. Værtens væv er stærkt opsplittet af snylterens intrusive organ. Røde pile markerer, xylem til xylem kontakt. Gule pile markerer parasit phloem (sivæv) med protein inklusioner omgivet af værtsceller, og mindst én af phloem cellerne indgår i interface langs phloemet i værtens ledningsvæv. Orobanche haustorier er dermed lige så avancerede som Cuscuta haustorier, og det er de to eneste slægter, hvor phloem til phloem kontakt er konstateret. Hos Orobanche crenata går phloem kontinuiteten endda et skridt videre end hos Cuscuta, idet der udvikles siporer ved ekspansion af plasmodesmer i grænsefladen mellem værts- og parasit phloem. Overførsel af mærkede kulhydrater og af et phloem-mobilt herbicid, glyphosat, har bekræftet, at sivævsporerne er funktionelle. Den veludviklede phloem forbindelse er medvirkende til gyvelkvæleres hurtige vækst. Klik på højre foto for at se plasmodesmer (hos Darlingtonia) i tværsnit og længdesnit. Plasmodesmers centrale rør er en forlængelse af det endoplasmatiske reticulum på tværs af cellevæggen og har en diameter på 10-15 nm. Det tillader ioner, sukkerarter og aminosyrer at passere, men efter en udvidelse til siporer kan større molekyler passere. Der går 1 million nanometer på 1 mm.


Orobanche er familiens største slægt med omkring 150 arter, der er vidt udbredte. Nogle af arterne er også introduceret langt fra deres primære udbredelsesområde. Eksempler er O. ramosa (t.v. Foto: C. Gracie) og O. minor (t.h. Foto C. Gracie), som er introducerede ukrudtsarter i det sydlige Afrika og USA. Sidstnævnte er også introduceret i Chile, New Zealand, og den er naturaliseret i Australien. Flere forskere foretrækker at opdele Orobanche i en række mindre, nært stående slægter, hvor kun Phelipanche skal nævnes her. De fleste arter er énårige, men ikke alle er undersøgt.



Hos mange arter ser blomsterne stort set ens ud som hos Orobanche picrides (t.v. Foto: G. Moreno Mor), men bortset fra farvevariationer er der variation i længden af det let buede kronrør og dets åbenhed. Nogle amerikanske arter har lange kronrør som O. fasciculata (Fig. i midten: Foto: R. Parker) og O. californica (t.h.). Næsten hele exofytten, inklusive højblade, bægerblade og dele af kronbladene, er dækket af kirtelhår (ses på O. picrides). Disse hår kan tjene til at holde myrer væk fra blomstens reproduktive dele. Bestøvning sker ved flyvende insekter, primært bier og hvepse. Der er to frugtblade, der danner en kapsel med ét rum. De små frø er talrige og tæller mere end 100.000 pr. plante for Orobanche minor. De naturlige amerikanske arter er ikke et problem på kulturplanter. Tilsvarende gælder i det nordlige Europa, men i Middelhavslandene forårsager 6 arter udbytte nedgang i solsikke, tomat, kartoffel, tobak og bælgfrugter, og disse arter har en tendens til at brede sig.


De fleste holoparasitter er flerårige, men de mest skadelige er étårige. Orobanche cumana, der angriber solsikke, er dog flerårig. Udbredelsen af Orobanche er i Nordamerika, omkring Middelhavet og østpå til Kina. O. minor og O. ramosa (begge vist ovenfor) er som nævnt blevet introduceret i flere lande rundt om i verden. O. ramosa ankom til Texas så sent som i 2000. Spredning er i de fleste tilfælde sket med ikke ordentligt rensede frø af kulturplanter. Orobanche angriber primært tokimbladede planter fra familierne Solanaceae, Fabaceae, Cucurbitaceae, Asteraceae, Apiaceae, Brassicaceae og hamp (Cannabis sativa) fra Cannabaceae. Adskillige andre afgrøder parasiteres af Orobanche arter ud over dem, der er anført i tabellen, og en håndfuld andre Orobanche arter end de anførte forårsager problemer lokalt. Værtsspecificiteten varierer betydeligt, hvor Orobanche cumana (Fig. t.v. Foto: www) er den mest værtsspecifikke, da den sjældent ses på andre værter end solsikke. Bemærk at solsikke bladene hænger som tegn på vandstres. Udbyttetabet er praktisk talt lige så alvorligt som ved skader forårsaget af Striga. Mere end en million hektar solsikke har været påvirket af O. cumana i Rusland, men den nuværende situation er ukendt. I Spanien blev 20.000 hektar påvirket i 1990’erne, og i Tyrkiet et ti gange så stort område. Økonomisk set er de største tab forårsaget af Orobanche spp. på hestebønner, linse, tobak og tomat. Ifølge et estimat forårsager O. ramosa et udbyttetab på 33% hos tobak, hvis hver værtsplante ernærer 1,5 parasit. En af de mest skadelige med mange værter er Orobanche crenata, (Fig. t.h. Foto: D. M. Joel) Bemærk, at den ikke inficerede gulerod i midten har en meget kraftigere rod end de inficerede. O. crenata kan forårsage død hos hestebønne (Vicia faba). I Spanien, Portugal, Marokko og Syrien er det blevet anslået, at op til 63% af det beplantede areal eller 200.000 ha er angrebet.


Phelipanche er en af de slægter, der er udskilt fra Orobanche, og det er ikke klart, hvor mange arter der er i slægten. Den middelhavske P. lavendulacea (Fig. t.v. Foto: G. Moreno Mor) har bevæget sig rundt i det taksonomiske system. Arten forekommer på dyrket ord, men er relativt harmløs, mens P. aegyptiaca angriber en lang række kulturplanter med udbytte tab som følge. Aeginetia er en anden lille slægt bestående af tre arter. Dens primære udbredelse er i Østasien, men ligesom andre parasitter, der angriber afgrøder, er den blevet introduceret i adskillige lande uden for dens naturlige udbredelse. Der er ingen synlige stængler, og de enkelte blomster optræder på lange stilke som hos Orobanche uniflora. Bægerbladet er ret dominerende, men den ydre del af blomsterkronen træder frem gennem en spalte på den ene side af bægerbladet som hos A. indica (Fig. t.h. Foto: D. M. Joel). En kapsel kan producere op til 70.000 frø. Den er en trussel mod majs, ris og sukkerrør i Indien og SØ-Asien.


To yderligere holoparasitiske slægter, Aeginetia, og Christisonia, omfatter arter, der angriber majs, ris og sukkerrør i Syd- og Østasien. Aeginetia indica (Fig. t.v. Foto: H Bänziger) synes at være den mest skadelige af de tre arter. Blomsterfarven varierer betydeligt, som det fremgår af de viste fotos. I naturen er A. indica normalt parasit på græsser af slægten Miscanthus. En beslægtet mindre art med en gul læbe, A. pedunculata, er for nylig rapporteret at parasitere sukkerrør i Bengal-provinsen i Indien. Sukroseindholdet i rørsaft fra ikke inficerede planter er i gennemsnit 16,3%, men kun 2% i inficerede planter, og tabet er 100%, når parasitten får sukkerrørene til at visne før høst. I Christisonia er C. wightii den mest problematiske, men den angriber kun sukkerrør på Filippinerne, mens arten på Fig. t.h. C. siamensis (Foto: H. Bänziger) tilsyneladende kun har vilde værtsplanter. Stilkene er mindre end 3 cm høje, så kun de smukke blomster viser sig lige over jorden.
Bekæmpelse af rodparasitter
Betydelige problemer for landbruget i Spanien med Orobanche crenata var den direkte anledning til det første internationale symposium om parasitiske ukrudtsplanter, der blev afholdt i 1973 på Malta. Hvert tredje eller fjerde år siden da har forskere mødtes i et internationalt forum for at diskutere parasitiske planters biologi, og hvordan de skadelige kan bekæmpes. Der er gjort en stor indsats for at identificere spiringsstimulerende stoffer, teste resistens og virkningerne af herbicider, forstå spiringens fysiologi, haustorie anlæggelse og forskellige aspekter af parasit-vært-fysiologi samt avle resistente sorter af værtsarter. Der er opnået lovende resultater, men de mest effektive bekæmpelsesmetoder er ikke nødvendigvis anvendelige overalt af praktiske eller økonomiske årsager, og generelt kan det fastslås, at forebyggelse er bedre en bekæmpelse efter, at angreb har slået an. Som forebyggelse kan bl.a. anbefales brug af certificerede frø, anvende resistente sorter, vekseldrift med ikke brugbare værter, dybdepløjning og rengøring af redskaber.

Manuel udryddelse af Striga hermonthica på denne Sorghum mark er nytteløst m.h.t. at redde udbytte, men det er nyttigt at fjerne blomsterne, så parasitten ikke sætter frø. Lugning er i mange afrikanske og asiatiske lande normalt et kvindearbejde. Det påstås, at mændene mister potensen, hvis de foretager lugearbejde. Orobanche frø bevarer spireevner i mere end 10 år og Striga arter i mere end 20 år. Foto: D. C. H. Plowes).
Dybdepløjning for at begrave parasitfrøene godt under afgrødernes rodsystem og spirings stimulanternes virkningszone kan være gavnligt, men ikke altid praktisk muligt. Dette skyldes et tyndt jordlag og/eller dyrt maskineri. Dyrkning og lugning på små marker er måske ikke dyrt, men gentagen dyrkning i årene efter dybdepløjning kan bringe levedygtige frø tættere på overfladen igen og forårsage genangreb (Foto: www).


Fattig landsby nær Kisumu, Kenya besøgt i 1990, hvor afmagret kvæg græssede på udtørret jord omgivet af majsmarker angrebet af Striga hermonthica. Rengøring af værktøj og sko efter markarbejde er vigtigt, og kvæg bør ikke flyttes mellem angrebne og rene marker. Kvæg kan dog være til en vis hjælp, når de græsser på angrebne marker. I områder med nomader har geder og kvæg tidligere holdt Striga-angrebet på et lavere niveau.


Afbrænding vil forhindre spredning af frø, men det ødelægger også afgrøden, og nye parasitplanter vil blive rekrutteret fra frøbanken (de levedygtige frø i jorden) i mange år fremover. Afbrænding har også den effekt, at asken forsyner jorden med flere mineraler, og bedre voksende og sundere værter er til fordel for parasitterne, som derefter får en bedre frøsætning. Brande giver også lysninger og forbedrede lysforhold for spiring af både rodparasitter og potentielle værter. På billedet t.v. er afbrænding brugt som naturpleje i Kruger Nationalpark. Solarisation er en metode til at lade solstråling øge jordtemperaturen så meget, at frøene dræbes, og frøbanken reduceres betydeligt. Jorden dækkes med klar polyethylenplast (Foto: www) på solrige dage, og den bedste effekt opnås, når jorden er våd. Metoden er også effektiv mod skadedyr i jorden og andre ukrudtsfrø.
Brug af braklægning har fordele og ulemper. Frøproduktionen kan være lavere efter braklægning på en bestemt mark, men parasitterne bruger mange vilde arter som værter samt planter fra resterende frø fra afgrøder. Braklægningen bliver derfor ofte en kilde til spredning af frø til dyrkede marker. Gødskning kan øge udbyttet selv i områder inficeret med Striga og Orobanche. Op til en vis grænse drager parasitterne fordel af velnærede værtsplanter, men over dette punkt reduceres parasitternes vækst og frøsætning. Dette har vist sig især sandt for kvælstofgødskning. De vilde stammer af Striga og Orobanche er tilpasset til at trives i relativt magre jorde, og eksperimenter med tilsætning af ammoniumbaseret kvælstof har reduceret og forsinket spiring af Striga asiatica og har reduceret levedygtigheden af frø hos både Striga og Orobanche. Nitratbaseret gødning synes ikke at have en lignende effekt. Eksperimenter fortsætter med at finde den optimale sammensætning og koncentration af den gødning, der skal anvendes i håb om, at værdien af det forbedrede udbytte vil overstige omkostningerne ved at bruge gødning.

Sædskifte anbefales af en række årsager, men er ofte af lav værdi i forhold til parasitiske ukrudtsplanter. Det skyldes den lange levedygtighed af frø, der er lagret i jorden (frøbanken). I afrikanske regioner, hvor parasitterne kommer ud af kontrol, forlades angrebne marker typisk efter den samme type afgrøde er dyrket i 4-5 år, og nyt land tages under dyrkning. Det betyder ofte afbrænding af skovområder, en procedure, der har mange alvorlige langsigtede konsekvenser for økosystemer og klima, udover det faktum, at de forladte marker er kilder til mere frøspredning fra parasitter. Sædskifte med fældeværter (falske værtsplanter), der inducerer spiring af parasitten, men ikke fungerer som vært, vil dog bidrage til at reducere frøbanken. Bomuld (Gossypium) stimulerer spiring hos Striga, og hør (Linum usitatissimum, foto herover) stimulerer op til 30% hos Orobanche, men begge fælde værter modstår angreb.


Samdyrkning med fældeafgrøder kan også anbefales for at reducere frøbanken. Det reducerer naturligvis også udbyttet pr. hektar at dyrke to forskellige afgrøder side om side, når den primære anvendelse af den sekundære afgrøde er at fange parasitten. Fælde afgrøden skal naturligvis høstes, før parasittens frø er klar til spredning. Fælde afgrøder kan også være en del af sædskiftet. Sædskifte og samdyrkning med korn afgrøder (durra, hirse osv.) og bælgfrugter (ko-ærter, jordnødder osv.) anvendes ofte, men har ringe effekt og kan forværre situationen, hvis f.eks. sojabønne bruges som fældeafgrøde for Striga hermonthica (Fig. t.v. Foto: D. C. H. Plowes) i et område, der også er inficeret med Alectra arter. Det vil føre til en ophobning af Alectra. Samdyrkning med Desmodium arter (Fabaceae) kan dog vise sig at være nyttigt, da majs så viser resistens over for Striga. Et rodexudat, især flavonoider og isoflavonoider, fra Desmodium uncinatum (Fig. t.h. Foto: R. J. Augustin), er også identificeret. Rodexudatet efterligner de stoffer (strigolactoner), som værtsplanter normalt udsender for at aktivere Striga frøets spiring, men kimroden dør hurtigt, når den ikke finder en vært. Effektive fældearter for Striga, der anvendes i rotation med afgrøder, er vilde græsser som Rotboelia cochinchinensis, Pennisetum purpureum, Digitaria ciliaris, Andropogon gayanus og arter af Brachiaria og Setaria, men den negative side er, at et års udbytte går tabt. Sen såning er en fordel, hvis en tidligt modnende variant af afgrøden er tilgængelig. Så er det muligt at høste, før parasittens frø er modne. Der vil stadig være et reduceret udbytte, men parasittens frøbank reduceres.

Resistens over for Striga hermonthica i ris. A-C, Tværsnit af værtsrod med haustorium (P). A, Modtagelig kultivar Kasalath 9 dage efter spiring af Striga. Det intrusive organ (E) har passeret endodermis. B, Endofyttens intrusive organ er vokset rundt om endodermis af den resistente kultivar Nipponbare 21 dage efter spiring. C, Udtømt værtsrod af Kasalath 25 dage efter spiring. H, hyalint body. Bar: 0,1 mm. – Efter Gurney et al. 2006.
Resistente sorter af afgrøden er uden tvivl den bedste løsning, men sådanne sorter er ikke altid tilgængelige, og hvis de findes, kan de være for dyre for mange landmænd. Mekanisk forsvar, f.eks. ved sklerenchym, som forhindrer haustoriet i at etablere sig, er omtalt tidligere. Det forekommer kun hos et par arter, men synes at være effektivt hos ris mod Cuscuta campestris i Indien. Af en noget lignende grund er ris kultivaren ’Nipponbare’ resistent over for Striga hermonthica (Fig. t.v.) Det primære haustorium får lov til at sætte sig fast, og det intrusive organ får lov til at trænge ind i værtens cortex. Endodermis, der omkranser ledningsvævet, gennembrydes dog sjældent, og endofytten (den del af snylteren der er inde i værten, modsat exofytten) fortsætter med at vokse rundt om ledningsvævet uden at være i stand til at etablere xylem-til-xylem-kontakt, hvilket er afgørende for parasittens overlevelse. Hvad der får endodermis til at afstøde endofytten i den resistente variant, er endnu ikke forstået, men ved hjælp af tilbagekrydsnings indavl mellem resistente og modtagelige forældre er der identificeret vigtige gener, som kan være nyttige i avlsprogrammer for at forbedre værtens resistens.


Der findes flere former for resistens, som kan være effektive på forskellige stadier af parasittens livscyklus. Der er også forskellige måder at opnå resistens på, herunder genteknologi. Udvælgelse og avl af resistente sorter har i et vist omfang været succesfuld i adskillige afgrøder, såsom tomat mod Phelipanche aegyptiaca og Striga gesnerioides. På Fig. t.v. har P. aegyptiaca i forgrunden angrebet en ikke resistent sort af tomat og i baggrunden en resistent sort på en forsøgsmark i Israel (Foto: D. M. Joel). T.h. ses vignabønner (Vigna anguiculata) angrebet af Alectra vogelii på forsøgsmark i Namibia (Foto: A. Larsen). De to rækker t.h. er en resistent sort, mens de øvrige rækker er en ikke-resistent sort. Billedet er taget efter høst, så vignabønnerne er visne. Man har også resistente sorter af durra (Sorghum) mod Striga hermonthica og S. asiatica. Sorter, der viser resistens, kendes også hos solsikke og hestebønner, mens der kun har været ringe succes med at finde resistente stammer i hirse. Den resistens, der findes i ovenstående sorter, er ikke total, men resistente sorter angribes sjældnere, og de har et forbedret udbytte sammenlignet med ikke-resistente sorter. Nogle sorter kan udvise tolerance, hvilket betyder, at værten fungerer normalt på trods af at være parasiteret. Når en resistent sort findes, skal den testes i marken, og hvis den lykkes, skal den dyrkes i stor skala for at høste såsæd. Det er dyrt, og mange mindre samfund har ikke økonomi til at købe såsæden uden tilskud.

Biologisk bekæmpelse har åbenlyse fordele, selvom bekæmpelsen sjældent er fuldstændig. Kun få metoder er blevet afprøvet mod parasitiske planter. Herbivori med insekter som fluen Phytomyza orobanchia (Diptera, Agromyzidae) har fungeret. Den er kendt for at bruge 21 arter af Orobanche som værtsplanter for sine larver. Larverne æder sig gennem stængler og kapsler og spiser så mange af frøene, at frøproduktionen reduceres med 30-80% (Fig. herover. Foto: K-H Linke et al. 1989). A, nedre stængel mineret af larver. B, voksen flue. C, kapsel med larve (rød markering) og devist spiste frø. Fluen har naturlige fjender og formerer sig ikke tilstrækkeligt i koldt vejr, men da 500-1000 voksne fluer/ha blev sat fri i det tidligere Sovjetunionen i begyndelsen af Orobanches fremkomst over jorden, opnåede man en reduktion i Orobanche frøproduktionen på op til 96%. Behandlingen skal gentages over flere år på grund af frøenes holdbarhed på 10-15 år, og metoden har kun potentiale inden for fluens naturlige område. – Svampe kan også bruges til biologisk bekæmpelse. Fusarium oxysporum og F. arthrosporioides forårsager rodråd i f.eks. Orobanche cumana, der snylter på solsikke. Effekten er god i laboratoriet, men det er en anden opgave at gøre metoden effektiv og økonomisk til feltbrug. Myceliet skal opbevares levende i store mængder og senere fordeles over store områder. Genteknologi er i gang for at øge svampenes virulens, dvs. øge deres evne til at forårsage sygdomme hos Orobanche. Fusarium reducerer også spireevnen hos Striga.

Rodparasitter i landbruget kan også bekæmpes med kemikalier, og brug af kemiske spiringsstimulatorer som ethylen og strigol har som omtalt tidligere været ret succesfuld på Striga, men metoderne er dyre og indtil videre primært for højt udviklede lande. Ideen er at stimulere parasitfrøene til at spire på et tidspunkt, hvor der ikke er nogen værter tilgængelige. Efter introduktionen af Striga asiatica (t.v.) i North og South Carolina er ethylen blevet brugt som spiringsstimulerende middel. 1,6 kg/ha blev injiceret i jorden på et tidspunkt, hvor frøene er forkonditionerede og klar til at spire. Gassen diffunderer gennem jorden i mere end 1 m fra injektionsstedet, og 90% af de forkonditionerede frø spirede og døde kort efter. Ethylen er også effektivt mod S. hermon-thica, men har ingen effekt på S. gesnerioides og Alectra. Nogle af de kunstige stimulatorer, der ligner strigols kemiske struktur, er dog effektive, men hverken disse eller ethylen har praktisk værdi i udviklingslande på grund af de høje produktions- og distributions omkostninger.
T.v. Striga asiatica på majs. Foto: D. C. H. Plowes.
Jordfumigering er en metode til at desinficere jorden for levende parasitfrø samt diverse skadedyr. Det mest effektive kemikalie er methylbromid (brommethan), som er brugt mod Orobanche. De negative aspekter er dog betydelige. Kemikaliet er giftigt, det kan kun transporteres i tryktanke, der kræves specielt udstyr til distribution, og en gastæt afdækning af det behandlede område er nødvendig samt store mængder af fumiganten (300-400 kg/ha). Metoden er tydeligvis meget dyr, men kan have værdi til rengøring af redskaber og opbevaringsfaciliteter m.m. Frøbehandling af korn anvendes, men har ringe effekt. Nogle landmænd i Nigeria lægger deres såsæd i blød i saltlage eller et ekstrakt af Parkia filicoidea (Fabaceae) og hævder, at det reducerer Striga infektion. Brugen af herbicider er altid tvivlsom på grund af generel toksicitet og manglen på undersøgelser af langtids miljøeffekter. Der kræves også specielt udstyr, teknologi og træning til deres anvendelse; derfor er metoderne dyre og ikke egnede i udviklingslande.
Marker inficeret med Striga asiatica, S. aspera og S. hermonthica kan behandles før såning ved at påføre trifluralin (2,6-dinitro-N,N-dipropyl-4-(trifluormethyl)benzenamin) overfladisk i jorden, men metoden er meget arbejdskrævende bl.a. fordi majs- eller durra frø skal plantes i hånden og placeres under zonen med herbicidet. Når kimplanterne har nået en højde på 20 cm trækkes den behandlede jord op omkring stilken, og antallet af Striga planter reduceres betydeligt. Andre herbicider anvendes også før såning/plantning, men en god effekt afhænger af nedbør for at vaske herbicidet ned i jorden. Siden 2005 har man haft frø af en naturlig mutant af majs, der er resistent over for det systemiske herbicid Imazapyr. Frøene bades i Imazapyr, som dræber den spirede S. asiatica, men ikke den resistente vært, og kombineret med gødskning er bekæmpelsesmetoden lovende.
Andre systemiske herbicider såsom dicamba (3,6-dichlor-2-methoxy-benzoesyre) påføres majs- og durrablade cirka 35 dage efter vækst over jorden. Herbicidet transporteres så til rødderne, mens planterne vokser. Timingen er vigtig, og sprøjtning skal ske i fasen, hvor parasitten anlægger haustorium, men mindst to uger før afgrøden begynder sin reproduktionsfase. Herbicidet bekæmper også mange konkurrerende ukrudtsarter, men i afgrøderne er det kun effektivt i cirka to uger. Herbicider til direkte sprøjtning på parasitten er også tilgængelige og effektive, men de dræber kun de sprøjtede planter og forhindrer ikke nye i at spire frem. Brug af herbicider mod Orobanche har ikke vist lige så stor effekt som mod Striga. Nogle systemiske herbicider, såsom glyphosat og imazaquin, har været i brug på hestebønner, ærter og linse, men resultaterne har været inkonsistente. Herbicider mod Striga gesneroides og Alectra spp. synes endnu ikke at være tilgængelige.
{kind=link}
Afslutningsvis ser det ud til, at ingen af ovenstående kontrolmetoder er effektive eller praktisk anvendelige som den eneste metode. Derfor har integrerede kontrolsystemer, hvilket betyder brugen af flere af ovenstående metoder samtidigt eller successivt på samme afgrøde i nogen tid været nøgleordet for bekæmpelse af rodparasitter i udviklingslande. Genetisk baseret resistens hos værterne er fortsat en af de mest ønskelige komponenter i en integreret bekæmpelsesstrategi. De grundlæggende mekanismer, der styrer resistens over for parasitiske planter, er dog langt fra fuldt ud forstået. Udover fortsat forskning er et andet punkt vigtigt, information og undervisning. Landmændene såvel som andre har brug for at lære de parasitiske arter at kende såvel som deres livscyklus. Der gøres også en indsats i den retning, og plakater som på figuren (klik for forstørrelse) er opsat i afrikanske lande i Sahel-regionen, hvor folk samles. De ser måske enkle ud, men er et nyttigt initiativ.
Parasitter i frugtplantager og skovbrug

Med hensyn til skadelige parasitter på træer og buske er de fleste hemiparasitiske stængelparasitter fra Loran-thaceae og Viscaceae, men også nogle rodparasitter har træer som vært. Det gælder Osyris, en slægt i Santalaceae med 3-4 arter, der findes fra Portugal til Indien. Fra Israel er det rapporteret, at O. alba (Fig. t.v.) angriber afgrøder i frugtplantager med mandel, pære og vin. Som det ses af billedet, har Osyris både hun- og hanplanter.
Blandt Orobanchaceae er rodparasitten Seymeria cassioides en hemiparasit, der forårsager problemer i skovbruget. Den angriber fyrretræer i det amerikanske syd. Hos Balano-phoraceae udvikler holoparasitten Thonningia sanguinea (Fig. t.h. med talrige røde hunblomster. Foto: Y. Caraglio) tyloser i vævet hos gummitræet Hevea brasiliensis. Tyloser er udposninger fra parenchymceller, der vokser ind i værtens xylem og blokerer for vandtransporten. Værtens væv trykkes også sammen, og xylemet splittes op i strenge, der gør veddet svagere. Thonningia anses derfor for at være farlig for gummiplantager. Thonningia er en monotypisk afrikansk slægt med hun- og hanblomster.



Arceuthobium med ca. 42 arter er den mest avancerede slægt i Viscaceae, hvor langt de fleste arter hører hjemme i Nordamerika. Der er en enkelt art A. oxycedri i Europa, og den når helt til Himalaya. Det er meget små hemiparasitiske stængelparasitter med hun- og hanplanter, der har meget stor effekt på værtens morfologi og anatomi. Herover vises to amerikanske arter t.v. A. camylopodum med næsten modne frugter på Abies magnifica (Foto: R Scharpf) og t.h. A. douglasii med hanblomster på Pseudotsuga menziesii.



Frugten er et énfrøet bær med viscinvæv. Fig. A viser næsten modne frugter af Arceuthobium campylopodum (Foto: R. Scharpf). Spredningen er unik i slægten ved at være eksplosivt selvspredende. Frugtfarven er grøn eller gulbrun, og sådanne farver tiltrækker ikke fugle. Eksplosions mekanismen er illustreret i Fig. B (baseret på Hawksworth hos King 1966). Før frugten er moden, bøjes stilken, så frugtens spids peger nedad. Når frugten er fuldt moden, udvikles der et enormt tryk på omkring 10 HPa i frugten. Til sidst kan det elastiske ydre lag ikke modstå trykket, stilken knækker i en forudbestemt zone med svage cellevægge (abscission zone), og frøet skydes ud som en kugle fra en kanon (Fig. C. Foto: Hawksworth). Hos A. americanum kan denne affyring forekomme for alle frø inden for to dage (normalt flere dage) i begyndelsen af september. Optagelser med et højhastighedskamera har vist, at starthastigheden er omkring 24 m/sek. Frøet følger en kurve, og den tilbagelagte vandrette afstand kan være op til 20 m eller i blæst meget længere. På den måde kan Arceuthobium arter rejse fra træ til træ og sprede sig i en skov. Én art, A. verticillatum, rapporteres dog at have ikke-eksplosive frugter, hvilket kan fortolkes som en evolutionær vending.



Naturligvis vil kun få procent af frøene lande til spiring på en egnet værtsgren, men at lande på en nål kan være lige så godt. Et sådant tilfælde er illustreret i figuren herover (Foto: R. Scharpf), hvor et frø af Arceuthobium abietinum er landet på en nål af Abies concolor. Den hygroskopiske viscin absorberer vand på en tåget eller regnfuld dag, og derefter glider frøet langsomt ned ad nålen, indtil det får kontakt med grenen. Ofte glider frøene også ned til den underste side af grenen, hvor de er bedre beskyttet. Så snart viscinen tørrer ud, mister den sine hygroskopiske egenskaber, og frøet sidder limet fast til grenen. Efter spiring og etablering af den unge endofyt følger en kortere eller længere periode, hvor parasitten kun består af endofytisk væv. I den periode er den usynlig udefra. Phoradendron californicum har også en sådan ’usynlig’ periode efter spiring, men situationen er kun generel hos Arceuthobium.

Endofytten fortsætter med at vokse dels som uniseriate filamenter (celler på række efter hinanden) og dels som multiseriate cortikale strenge. Disse strenge løber i cortex eller phloemet og producerer sænkere i værtens sekundære xylem. Efter nogle måneder, afhængigt af arten, produceres blomstrende skud fra de cortikale strenge. Samtlige exofyt skud af Arceuthobium campylopodum på Pinus contorta grenen t.v. repræsenterer således formentlig kun én plante.


Endofytten forårsager alvorlige ændringer i væksthormoners funktion hos værten. Der reageres på flere måder, men ofte ved at producere et stort antal mindre skud meget tæt på hinanden. Sådanne samlinger af uregelmæssige skud kaldes heksekoste og ses på figurerne herover af Arceuthobium douglasii på Douglas gran (Pseudotsuga menziesii). Heksekoste kan i høj grad reducere væksten og den kommercielle værdi af værtstræet. På figuren t.h. fra en bjergskråning i British Columbia er mindst tre træer døde. Visse svampe (Taphrina spp.) forårsager også dannelse af heksekoste.


Især to omstændigheder gør Arceuthobium til en frygtet parasit. For det første dens evne til at selvsprede sig via de eksplosive frugter og derved sprede sig gennem en skov uden at være afhængig af et spredningsmiddel. For det andet gør dens ofte meget lille størrelse det svært at opdage den, før det er for sent. Starten på en heksekost kan spores ved at se på værtstræets ændring i skudbygningen. T.v. ses et angreb af Arceuthobium campylopodum på lærk (Larix occidentalis). Værtens skudbygning består af langskud, hvorpå der sidder kortskud med nåle. Parasitten har forstyrret væksthormonerne i kortskuddet nærmest exofytten, så kortskuddet er blevet til et langskud (peger skråt opefter på billedet). Der vil herefter dannes flere langskud fra samme udgangspunkt, og der fremkommer en heksekost. Alle Arceuthobium arter forårsager skade på værten. Udover dannelsen af heksekoste forårsager parasitangrebet ofte hævelser i værtens grene, som det ses på figuren t.h. af A. campylopodum på Pinus contorta. Hævelserne kan føre til dannelse af revner og større åbne sår, som igen bruges til svampeangreb f.eks. infektion med Cytospora abietis, der forårsager trækræft. Andre problemer omfatter en generel svækkelse af træet, hvilket forårsager brud, tab af tømmer og gør rekreativ brug af skovene farlig. Nogle arter, såsom A. campylopodum, vil forårsage heksekoste hos én vært, men hævelser hos en anden værtsart.
De mest problematiske stængelparasitter tilhører uden tvivl slægten Arceuthobium, og området med den værste økonomiske påvirkning er det vestlige Nordamerika. Tabene forårsaget af de forskellige Arceuthobium arter beløber sig ifølge et estimat fra 1982 til 11,3 millioner m3 træ i det vestlige USA og 3,8 millioner m3 i det vestlige Canada, hvilket repræsenterer en værdi på flere milliarder dollars årligt, og disse estimater synes ikke at have ændret sig de seneste 40 år. Til disse tal kan lægges betydelige tab i Mexico, Guatemala, Belize, Den Dominikanske Republik, Indien, Pakistan og Kina.




Kommercielt dyrkede træer, der er hårdest beskadiget af forskellige vestlige Arceuthobium arter, er Pinus ponderosa, P. contorta (Fig. herover t.v. angrebet af A. americanum), P. jeffreyi, P. banksiana, Pseudotsuga menziesii (Fig. i midten angrebet af A. douglasii), Abies magnifica, A. concolor, Larix occidentalis (Fig. t.h. angrebet af A. campylopodum) og Tsuga heterophylla. I Lake District og det østlige Nordamerika er Pinus marina, P. glauca og P. rubens også alvorligt angrebet. Skaderne spænder fra reduceret vækst og frøproduktion til forvrængede værtsgrene, reduceret trækvalitet og til værtens død. Den tid, det tager at dræbe en vært, afhænger af en række faktorer, herunder den involverede art, infektionens sværhedsgrad, værtens styrke, økologiske og klimatiske faktorer og aktiviteten af skadedyr, især barkbiller (Scolytidae). I bestande med flere aldre er dødeligheden højest for træer med en diameter på mindre end 25 cm i bestande af Pinus ponderosa inficeret af Arceuthobium vaginatum ssp. cryptopodum og i bestande af Pseudotsuga menziesii inficeret af Arceuthobium douglasii. Når parasitterne har nået den øvre del af værtstræerne, reduceres væksten i både højde og tykkelse, og træerne begynder at dø (Fig. t.v. og herover i midten).


Hos Arceuthobium udviser adskillige arter høj værtspecificitet, dvs. de accepterer kun få værts arter. A. minutissímum (Fig. t.v. Foto: G. Glatzel), der er den mindste art, er kun registreret fra Pinus griffithii. A. oxycedri (Fig. t.h. Foto: G. Glatzel) kendt fra Europa har kun Juniperus spp. som vært i naturen, og A. douglasii, A. pusillum og A. americanum er stort set begrænset til henholdsvis nåletræerne Pseudotsuga, Picea og et par Pinus arter.
Tilsyneladende er Arceuthobium arter i stand til at konkurrere om de tilgængelige værtsarter. I Colorado Front Range er Pinus contorta den primære vært for A. americanum, og Pinus ponderosa er den primære vært for A. vaginatum ssp. cryptopodum. I bestande af Pinus ponderosa, hvor begge parasit arter er til stede, er kun 13 % af træerne inficeret med A. americanum, mens 64 % af træerne er inficeret i bestande uden A. vaginatum ssp. cryptopodum. Infektionsgraden er omvendt i en bestand af Pinus contorta. Lignende eksempler med andre arter er også kendt.
Arceuthobium snylter kun på nåletræer, men de fleste slægter omfatter adskillige arter, der ikke er blevet bemærket som værter. I USA er der f.eks. 3 arter af Picea, 4 af Abies, 14 arter af Pinus foruden Pseudotsuga macrocarpa, som ikke angribes. Det kan dog ikke konkluderes, at disse arter alle er resistente over for Arceuthobium, da det naturlige udbredelsesområde kan være uden for udbredelsen af nogle arter af Arceuthobium.
Hvis en parasit har succes med at etablere sig på en vært, skal der være vævskompatibilitet mellem de to parter. Normalt accepteres et stort antal tilsyneladende egnede planter, der forekommer inden for en bestemt parasits’ udbredelsesområde, ikke som værter for den pågældende parasit. I de fleste tilfælde vil forklaringen være en form for biokemisk/fysiologisk inkompatibilitet. Det betyder, at værtsgenkendelsen er negativ eller ikke-funktionel, og at de ikke-parasitiske planter har et effektivt biokemisk forsvar. Et eksempel på biokemisk inkompatibilitet ses hos Arceuthobium, som aldrig er blevet fundet på tokimbladede træer. Der findes også eksempler på mekanisk forsvar, hvor f.ks. korkdannelse forhindrer parasitten i trænge ind, men det er ikke konstateret hos Arceuthobium.

Stængel parasitterne Cassytha og Cuscuta er tidligere omtalt som skadevoldere i landbrug og gartnerier, men især Cassytha kan også skade træer, og den udbredte C. filiformis forårsager problemer ved angreb i avocadoplantager (Persea americana), f.eks. i Australien. Her skal det handle om stængelparasitter fra Loranthaceae, der er den største familie i Santalales med 73 slægter og lidt over 900 arter. Loranthacéer kaldes i engelsk litteratur for de prangende misteltener (showy misteltoes) på grund af de ofte spektakulære røde og gule blomster. Figuren herover viser Psittacanthus ramiflorus med lange rørformede blomster, der anlægges direkte fra grenen. (Foto: M. Bonifacino). Loranthacé blomster kan blive mere end 20 cm lange. Det står i modsætning til de meget små blomster i Viscaceae. Desværre er mistelten den fælles betegnelse for alle stedsegrønne parasitter i familierne Loranthaceae, Eremolepi-daceae og Viscaceae, som kun har det til fælles, at de lever som hemiparasitter på forskellige værtstræer. I mange europæiske lande refererer mistelten kun til arter i Viscaceae. Der er således en åbenlys grund til at bruge de latinske eller videnskabelige navne så meget som muligt. Loranthaceae er hovedsageligt subtropisk og tropisk, men undgår de tørreste områder. Derudover er der en isoleret forekomst af Loranthus europaeus i det tempererede Mellem- og Sydøsteuropa.



Der er tre rodparasitiske slægter i Loranthaceae, der kun etablerer sig v.hj.a. sekundære haustorier. Alle øvrige Lorantha-céer anlægger først et ofte ret stort primært haustorium, som de mest avancerede (f.eks. Psittacanthus) er fuldstændigt afhængige af, mens mange andre også danner sekundære haustorier fra epicortikale rødder. Øverst ses en epicortikal rod af Alepis flavida med fire sekundære haustorier på Nothofagus solandri (underst) fra New Zealand (Foto: C. L. Calvin & C. A. Wilson). T.v. et primært haustorium af australske Amyema miquelii, hvor værten har dannet en krave kaldet en wood rose omkring haustoriet (Foto: G. Glatzel). T.h. ses Notanthera heterophylla fra Chile med unge bladskud vokse frem over sekundære haustorier fra epicortikale rødder (Foto: J. Kuijt).


Loranthacéer har mere komplicerede haustorier end vist øverst på siden. T.v. er alt ikke forveddet væv fjernet, så kun xylemet ses hos Psittacanthus ramiflorus på eg (Quercus sp.). Parasitten har sekundær tykkelsesvækst, og den seneste årring slutter ved pilen. Værtens delta-lignende wood rose xylem ses nederst (Foto: Kuijt & Lye 2005). Herover, det lyse væv er parasit væv fra Scurrula elata overvokset af værtens sekundære tykkelsesvækst (Foto: G. Glatzel). P, parasit. V, vært.


Nogle få Loranthacé slægter har små blomster med 4-7 frie blosterblade og insektbestøvning, men de fleste har relativt store prangende blomster med et nektarie og mere eller mindre rørformet sammenvoksede blosterblade i røde og gule farver, hvilket peger på fuglebestøvning. Støvdragerne er krøllet sammen i knoppen og sidder på indersiden af kronrøret (t.h.), ofte på en sådan måde, at blomsterne åbner sig ved eksplosion efter stimulering af næbbet på en besøgende fugl. Den eksplosive åbning forårsager en overpudring af fuglen med pollen. Blomsterne signalerer på forskellig vis, at de er klar til bestøvning. Hos Globimetula mweroensis (t.v.) er et farveskift fra rød til sort af kronrørets opsvulmede spids signalet til solfugle om, at blomsten er klar til at eksplodere ved berøring (Foto: D. C. H. Plowes). Hos nogle slægter er blomsten ventileret, hvilket betyder, at den før eksplosionen åbner slidser svarende til antallet af sammenvoksede blosterblade som hos Englerina inaequilatera (t.h. Foto: Polhill & Wiens 1998).



Agelanthus gracilis er en anden Sydafrikansk art, hvor slægten har ventilerede eksplosive blomster, der åbnes ved berøring med solfugles næb. Mens hos nogle slægter alle slidser skilles ved eksplosionen, så pollen spredes til alle sider, så er eksplosionen hos Agelanthus retningsbestemt, idet kun slidsen nærmest fuglens næb splitter helt, så pollenkornene slynges direkte i hovedet på solfuglen.


Det er ofte de samme fugle, der bestøver blomsterne og spreder de bæragtige frugter med blot et enkelt frø. Herover ses frugter af Erianthemum ngamicum, og en Golden breasted bunting (Emberiza flaviventris) har plukket et bær og er i færd med at klemme frø og frugtkød ud af bærret for at sluge det. Nogle fugle går kun efter frugtkødet og smører bærret af på værtstræers grene, hvor viscinen klæber frøet fast. Andre sluger frøene og lader de fleste passere fordøjelseskanalen, så de kommer ud som rosarier (se Fuglespredning af mistelten).


Problemer med arter fra Loranthaceae kan være udtalte lokalt i Australien. En række Amyema arter parasiterer tømmer- og prydtræer. Belastningen af parasitter kan være imponerende, som det ses herover. Disse figurer illustrerer, at når et træ bliver inficeret med én stængelparasit, er risikoen for at få flere parasitter øget på grund af de frøspredende fugles adfærd. Figuren t.v. illustrerer også epiparasitisme, dvs. at en parasit etablerer sig på en anden parasit. Her er træet Melaleuca
bracteata angrebet af Amyema congener, der selv er parasiteret af Notothixos subaureus og Viscum articulatum (Foto: R. Kendall). Herover t.h. har Amyema bifurcatum angrebet en Corymbia sp. (Myrtaceae) med få synlige brede grønne blade (Foto: R. Kendall). Når parasitten dækker mere end 50% af kronen, er værtstræet i fare for ikke at overleve. Amyema congener, Muellerina eucalyptoides og Decaisnina hollrungii er blandt de Lorantacéer, der oftest angriber dyrkede og eksotiske træer i Australien.


Parasittens størrelse og vægt kan resultere i, at værtsgrenen ikke kan bære den, og risikoen øges, når flere parasitter sidder på samme gren, og hele værten kan eventuelt vælte i stormvejr. T.v. er det australske Amyema pendulum på Eucalyptus sp. (Foto: G. glatzel). Vægten af en enkelt plante kan blive over 10 kg. T.h. er det amerikanske Phoradendon californicum på Cercidium microphyllum (Foto: D. Leatherman). Den kan nå 1–3 m i diameter og op til 4 m i udstrakt størrelse, hvor vægten så kan blive over 30 kg. Her kan så også nævnes en positiv effekt ved store og tætte Loranthacéer, idet fugle her kan få mad (nektar, frugter), skygge og redesteder.



I Indien forårsager Dendrophthoe falcata (t.v. Foto: C. & D. Frith) enorm skade på teak (Tectona grandis), som er den vigtigste træart i landet. Op til 40% reduktion i årlig tilvækst af teak er registreret. Parasitten kan endda føre til døden af hele træer, og af en eller anden uforklarlig grund er dødeligheden af teak højere end hos flere andre kombinationer af tropiske træer og stængelparasitter. En årsag til D. falcata’s succes på teak kan være, at den er mere skyggetolerant end de fleste andre Lorantacéer. I gennemsnit modtager parasitten kun 40% af det lys, som værten modtager, og den vil overleve, selv når værtens blade blokerer 70% af den indfaldende solstråling.
Nogle af de større Loranthacéer gør stor skade i Vestafrika, og ikke mindst Tapinanthus bangwensis (Fig. i midten Foto: G. Glatzel). Den bruger en bred vifte af værter og er blevet invasiv i plantager, siden kakao blev introduceret som afgrøde i 1870’erne. Problemet steg med skovrydning og praksissen med at dyrke kakao uden skyggetræer. Det er eksperimentelt vist, at spiring af frø og etablering af kimplanter af den lysafhængige parasit er op til tre gange mere sandsynligt i uskyggede sammenlignet med skyggede kakaotræer. Phragmanthera capitata er også et problem i plantager med teak og gummi. Tilstedeværelsen af andre store arter af Tapinanthus, Agelanthus og Globimetula forværrer kun situationen. I tropisk Afrika kan Helixanthera mannii lokalt være et problem i plantager med avocado, kakao, citrus, kaffe og kapok. I Mellem- og Sydamerika er Struthanthus orbicularis et problem i citrusplantager.
Viscum arter gør kun moderat skade, men kan dog volde problemer i Afrika, hvilket V. menyharthii giver udtryk for ved topdød i et Ficus træ (Fig. t.h. Foto: D. C. H. Plowes).
L


I Sri Lanka parasiterer Dendrophthoe longiflora eksotiske frugttræer som Annona squamosa (Fig. t.v. Foto: G. Glatzel). I Himalaya (Annapurna-området) forårsager Scurrula elata reduceret vækst og undertiden visning af Rhododendron arboreum. I Amerika forårsager adskillige Phoradendron arter problemer ved angreb på både løvtræer som eg (Quercus) og i den nordlige del nåletræer fra Pinaceae og Cupressaceae. Fig. t.h. viser P. californicum på Acacia sp. I Mexico parasiterer P. rhipsalinum Taxodium nåletræer. Ved den sydlige rand af Grand Canyon rapporteres det, at P. divaricarum og P. juniperinum dræber arter af enebær (Juniperus).

I mange år var nedskæring af stængelparasitter den eneste metode til at kontrollere angreb fra arter af Loranthaceae og Viscaceae. Afhængigt af arten skal mere eller mindre af værtsgrenen også skæres af på grund af endofyttens vækst som cortikale strenge inde i grenen. Ellers vil der ofte opstå nye skud fra endofytten (den del af snylteren der er inde i værten modsat exofytten) eller fra hæfteskiven. Nedskæring af grene inficeret med Arceuthobium vil dog ofte være nytteløst, da endofytten hos en række arter når helt til værtens skudspidser som illustreret herover ved en anatomisk undersøgelse af endofytisk væv hos Arceuthobium douglasii i en skudspids af værtstræet Douglasgran (Pseudotsuga menziesii). A, Længdesnit gennem skudspids med unge nåle i vækst. B, Tolkning af et enkelt cortikalt snit, hvor parasitceller er farvet røde. C, Talrige cortikale snit skåret med 10 µm mellemrum lagt oven på hinanden. Omrids af snit tegnet grønt. Endofytiske celler er røde. D, Som C, men snittene ligger centralt omkring skuddets akse. Det er dermed vist, at parasitten ligger som en cylinder i værten, og at parasitceller når ud i nålene. Foto: D. Lye 2006, Annals of Botany 97.


Brugen af herbicider er afprøvet, men der findes meget få herbicider til et system, hvor både vært og parasit er tokimbladede. Herbicider, f.eks. 2,4-D dichlorphenoxyeddikesyre, er sprøjtet på bladene af forskellige parasitter eller injiceret i værtens stamme, men med inkonsistente resultater. Herbicider kan bruges til at bekæmpe Dendrophthoe falcata på teaktræ, men så er det tilrådeligt at bruge herbiciderne i værtstræets løvfældende fase. Mere end 60 forskellige herbicider er blevet afprøvet på Arceuthobium, men ingen kunne forhindre endofytten i at spire frem igen. Det mest effektive kemikalie anvendt på Arceuthobium er vækstregulatoren Florel, der indeholder 2-chlorethylfosforsyre. Det forårsager tidligt skudfald (abcission) hos adskillige Arceuthobium arter. Afhængigt af, hvor effektivt Florel anvendes, kan skud abcissionen være op til 100%. Hos f.eks. A. campylopodum sker genspiring fra endofytten dog hurtigt, men frøspredning kan forhindres i en periode på op til fire år afhængigt af kombinationen af parasit og værtsart. Gentagen anvendelse kan have negative virkninger på værten. Billederne viser A. campylopodum på Pinus contorta (t.v.) og Larix occidentalis (t.h.).


Biologisk bekæmpelse ved hjælp af planteædere kan have et potentiale i Australien. Her er Loranthacéer ikke aggressive i stabiliserede økosystemer med en forskelligartet fauna, men balancen ændrer sig, når mennesket blander sig. Der findes også planteædere på Arceuthobium, men da de har udviklet sig sammen med parasitten, er det ikke sandsynligt, at de er effektive til praktisk bekæmpelse i skovbruget. T.v. ses A. campylopodum, hvor skud på en lavtsiddende gren af Pinus contorta er spist af hjorte, men det har ikke betydning som bekæmpelse. Det er forsøgt med svampesygdomme hos Arceu-thobium. Svampen Caliciopsis (= Wallrothiella) arceuthobii ødelægger hunblomsterne hos visse forårsblomstrende arter såsom A. americanum og A. douglasii (Fig. t.h. Foto: R. Scharpf) og forhindrer derved yderligere spredning af parasitten, men et angrebet træ kan ikke reddes. Svampe som Colletotrichum gloeosporioides og Neonectria neomacrospora er brugt til biologisk bekæmpelse af Arceuthobium tsugense i Canada. Begge svampe er delvist effektive forudsat, at værtstræet behandles med sår for at lette sporernes spiring.
Ligesom for rodparasitter er udvælgelse af resistente sorter den optimale langsigtede løsning på problemer med Arceuthobium i skovbruget. Der er mange eksempler på overgange mellem resistens og modtagelighed hos en vært over for en parasit. Træer, der viser delvis resistens, bør vælges til videre avl. For eksempel viste nogle træer af Pinus ponderosa og P. jeffreyi fra Oregon og Californien sig at være delvist resistente over for Arceuthobium campylopodum. Når frøplanter eller podede prøver plantes i bestande af meget modtagelige træer, er infektionsgraden betydeligt mindre end på den modtagelige bestand, selv efter 20 år. Delvis resistens er også blevet fundet i andre værts-parasit kombinationer.
Kortsigtede løsninger er primært baseret på skovbrugsmetoder baseret på princippet om, at forebyggelse er mere effektivt end at fjerne parasitten eller genplante inficerede bevoksninger. Dette omfatter blandt andet regelmæssig inspektion af bevoksningerne, beskæring af inficerede grene eller fældning af træer så tidligt som muligt i nyligt inficerede bevoksninger, fældning af bånd bredere end 20 m mellem rene og inficerede bevoksninger. 20 m er normalt den maksimale afstand, som de eksplosive frugter af Arceuthobium kan skyde frøene. Spredning over lange afstande af fugle forekommer kun lejlighedsvis. Detaljerede forvaltningsplaner, vejledninger til beskæring og beskrivelser af de vigtigste Arceuthobium arter er tilgængelige fra USDA Forest Service.
Supplerende Litteratur:
Geils B. W., J. C. Tovar & B. Moody (eds.) 2002: Mistletoes of North American Conifers. U. S. Department of Agriculture. Forest Service. 123 pp.
Heide-Jørgensen H. S. 2008: Danske og nordiske snylteplanter. – with Figure legends in English. 40 pp. Download her. Hele artiklen er oversat til engelsk. Se abstract 66.
Heide-Jørgensen H. S. 2008: Parasitic flowering plants – Rigt illustreret populærvidenskabelig bog. Brill. 438 pp.
Heide-Jørgensen H. S. 2004: Værtsvalg, økonomisk betydning og evolution – (Snylteplanter IV). Naturens Verden 87 p 42-61. Se abstract 53-56 for hele serien om snylteplanter.
Joel D. M, J. Gressel & L Musselman (eds.) 2013: Parasitic Orobanchaceae – Parasitic mechanisms and control strategies. Springer – Introduktions kapitel af H. S. Heide-Jørgensen.
Kim S. K. (ed.) 1988: Combating Striga in Africa. International Institue of Tropical Agriculture. 145 pp.
Kuijt J. 1969: The Biology of Parasitic Flowering Plants. – Et klassisk videnskabeligt hovedværk.
Moss J. T. & R. Kendall 2016: The Mistletoes of Subtropical Queensland, New South Wales and Victoria. Butterfly & other Invertebrates Club Inc. 134 pp.
Polhill R & D. Wiens 1998: Mistletoes of Africa. The Royal Botanic Gardens, Kew. 370 pp.
Scharpf R. F. 1993: Diseases of Pacific Coast Conifers. U. S. Department of Agriculture. Handbook 521. Forest Service. 199 pp.
Watson, D. M. 2011: Mistletoes of Southern Australia. CSIRO Publ. 188 pp.
Der findes desuden enkelte artikler på dansk om nældesilke, skælrod og gyvelkvælere i Botanisk Forenings medlemsblad ‘Urt’.
Der er talrige indgange på internettet, hvor den mest omfattende er:
The parasitic plant connection.
H. S. Heide-Jørgensen, december 2025