Rundbladet soldug er den mest udbredte soldug art i Danmark. Den er cirkumpolar inklusive det sydlige Grønland. Den trives fint med konkurrence fra tørvemos (Sphagnum). På billedet th. vokser soldug tæt rundt om en grundvands regulere klitlavning i den rødbrune bræmme, hvor der hverken er for vådt eller tørt.
Drosera rotundifolia blomstrer i august. De 5-tallige blomster er kun åbne midt på dagen, og de sidder højt hævet over fælderne. I tørt og solrigt vejr kan slimen på tentaklerne tørre ud. På billedet herunder ses i midten et blad foldet sammen om et bytte som resultat af den på hovedsiden omtalte auxin regulerede vækstbevægelse.
Langbladet soldug - Drosera anglica
Langbladet soldug har i Danmark kun få voksesteder på højmoser og i fugtige klitheder i Jylland. Dens totale udbredelse er næsten cirkumpolar. Den findes dog ikke i Island og Grønland, men i Nordamerika når den op i Alaska. Der findes også nogle subtropiske forekomster på Hawaii og i det sydlige Europa og sydlige Japan. Den trives fint i sure omgivelser mellem Sphagnum. Den overvintrer ved vinterknopper kaldet hibernakler. Arten er selvbestøende og menes opstået som en diploid hybrid mellem den nordamerikanske D. linearis og D. rotundifolia. Drosera anglica x obovata er en steril hybrid med D. rotundifolia. Hybriden forveksles undertiden med Drosera intermedia.
Liden soldug - Drosera intermedia
Liden soldug findes især i moser, hvor den står under vådere forhold end de to øvrige danske arter, rundbladet og langbladet soldug. Den har hovedudbredelse i det vestlige Europa og østlige Nordamerika, men når ned i det nordlige Sydamerika. I tempereret klima overvintrer den ved hibernakler. Bladene er oprette, og formen er intermediær mellem de to førnævnte arter. Blomsterstanden virker sidestillet, idet den er bøjet til siden, mens den står opret centralt i rosetten hos de to andre danske arter. Bomstrer fra juni til august. På billedet t.h. vokser den sammen med blærerod Utricularia australis. – Drosera anglica x obovata er en steril hybrid med D. rotundifolia. Hybriden forveksles undertiden med Drosera intermedia.
Der er mindst 25.000 naturlige orkidé arter, og de er så godt som alle sammen insektbestøvede. Nogle kan bestøves af flere forskellige insekter, men mange er så specialiserede, at de kun kan bestøves af én bestem insekt art. Der er således sket en co-evolution mellem orkidé og insekt. Det sikrer en effektiv fremmedbestøvning, men øger risikoen for, at begge arter uddør, hvis livs-forholdene for bare den ene art ændrer sig i uheldig retning. Dette indlæg illustrerer en række af de mest raffinerede tilfælde af samspil mellem blomst og bestøver, men først skal orkidé-blomstens bygning forklares.
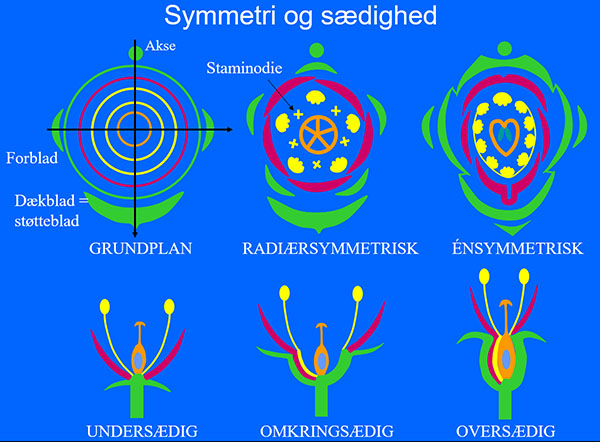
En blomst består af et antal bladkranse, inderst frugtblade, yderst bægerbade. Antallet af blade i hver krans varierer, og ofte er der kun én krans af støvblade, selv om der er vist to på grundplanen. Det er også almindeligt, at der sker reduktion af enkelte blade eller sammenvoksninger i en krans. Reducerede ikke funktionelle støvblade kaldes staminodier. Blomsternes sædighed er illustreret herover t.v. Orkidéer har oversædige blomster, dvs. at blosterbladene (bæger- og kronblade) og støvbladene er vokset sammen med frugtbladene næsten til spidsen af frugtbladene, og så er orkidé blomsten zygomorf, dvs den har kun et symmetriplan ligesom f.eks. ærteblomster. Udgangspunktet svarer til en 3-tallig liljeblomst, og orkidéerne og liljerne har derfor af mange været placeret i liljeordenen (Liliales). Mange orkidéer har en spore på det bagerste kronblad (petal), men mens knoppen udvikler sig snor frugtknude og blomsterstilk sig, så sporen i den udsprungne blomst vender væk fra blomsterstandes akse. Det kaldes, at blomsten er resupinat.
Det er nemt at se snoningen (resupinationen) hos Vanda, da frugtknuden er furet svarende til ryg- og bugsømme i de 3 frugtblade. Det er dog sådan, at frugtknuden regulerer snoningen, hvis blomsterstands aksen er skæv eller krummer, så blomsten som hovedregel orienteres med læben nederst, dvs. i tyngdekraftens retning. Det er formentlig den mest hensigtsmæssige position i relation til bestøveren, der normalt bruger læben som landingsplads.
Herover vises eksempler på variation i petaler og sepaler. Hos Vanilla imperialis (øverst t.v.) er de næsten ens, bortset fra læben, hvilket vel er det mest almindelige. Hos Masdevallia tovarensis (nederst t.v.) er sepalerne dominerende, og petaler og læbe er forsvindende små, mens de to laterale petaler hos Phragmipedium phillippinense er ekstremt lange, smalle og snoede, og læben er formet som en tøffel.
Eksempler på læbens variation. Phalaenopsis hybrid (t.v.) har stor læbe med vinger. Coelogyne lawrenciana (midten) har samme læbeform, men udstyret med papiller. Renanthera imschootiana (t.h.) har næsten ingen læbe.
Hos Stanhopea oculata (t.v.) er læben nærmest monstrøs med øjeplet og en gaffelformet afslutning. Aerangis biloba (t.h.) er eksempel på, at læben kan være udstyret med en spore, der indeholder nektar. Sporen er hos nogle arter blot en lille udposning, hos andre kan den være over 30 cm lang. Andre blosterblade kan også være udstyret med en spore.
Det nok mest specielle ved de fleste orkidé blomsters konstruktion er sammenvoksningen mellem det eneste udviklede støvblad og det nærmeste frugtblad til den såkaldte søjle/griffel-støtte. Samtidig er de to laterale frugtblades fang vokset sammen til et, mens det mediane støvfang danner en udvækst kaldet rostellum, der er et næb mellem støvfang og støv-knappen til forhindring af selvbestøvning. Spidsen af rostellum er forsynet med en klæbeskive (viscidium), der spiller en rolle, når pollinierne fra en anden blomst skal opsamles. Pollenkornene hos orkidéer er klæbet sammen i støvkøller (pollinier), der ses pillet ud af støvknappen på billedet t.h.
Zygopetalum mackay. T.v. peger pilen på støvknappen og rostellum. På det midterste foto er pollinierne blotlagt. Fanget sidder i en hulhed på søjlens underside. Fangets overflade producerer en sukkerholdig væske (glinser på billederne), som pollenkornene fanges og spirer i. T.h. ses en blomst, der er blevet bestøvet, og fangets overflade er tørret ud og har en brunlig farve. Støvknappen er tom.
Støvfanget kan være 3-delt eller 2-delt, men oftest er de to fang som her hos Rossioglossum grande vokset sammen til ét. Spidsen af rostellum er her næsten sort.
Skemaet angiver bestøvningsmåder hos blomsterplanter. Langt de fleste orkidéer har insektbestøvning, men fugle er også på listen. Selvbestøvning kan forekomme, og det er ikke ualmindeligt, at selvbestøvning træder til som en sidste udvej. Hvis blomsten ikke er blevet bestøvet ved et insekts medvirken, kan pollinierne kort før blomsten visner falde ud og svinge ned og ramme støvfanget, så der sker en selvbestøvning. Ophrys apifera er altid selvbestøvet og derfor ved en evolutionær endestation. Den er for få år siden fundet ved Vejle som ny art i Danmark. Foto: A Woodward, Flickr.
Hos orkidéer med hun- og hanplanter som hos Cycnoches chlorochilon kan selvbestøvning naturligvis ikke foregå. Hos denne art ligner de to køn hinanden meget, men hos andre arter er der stor forskel. Det er hun t.v. og han t.h. Bemærk den lange krumme griffelstøtte. Om bestøvning hos Cycnoches, se senere. Foto af hunblomst: J Pestana, Flickr.
Den tvekønnede blomst har effektiviseret insektbestøvningen, idet insektet både kan opsamle og aflevere pollen ved samme besøg. Ingen insektbestøvning uden, at der er noget at komme efter for insektet i form af føde, hvile/varme og gemmesteder eller æglægningspladser. T.v. har en humlebi fyldt pollenkurven på bagbenet under besøg hos en Cosmea, men den går ikke hos orkidéerne. Der er ikke råd til, at pollinierne bliver spist. Der er brug for alle pollen, for orkidéerne producerer mange meget små frø. Der er talt 3.770.000 frø/kapsel hos Cycnoches chlorochilum. Så orkidéerne må have andet at byde på, det vil først og fremmest sige nektar. Det kan dog give det problem, at orkidéer, der bruger pollensamlende insekter som bestøvere, indirekte bliver afhængige af, at der er andre blomster i nærheden, der er leveringsdygtige i pollen. T.h. ses Platanthera bifolia (Bakke-gøgelilje) med lange sporer fyldt med nektar.
Selv om nektar er vigtigste lokkemiddel, så regner man ikke med, at mere end to tredjedele af alle orkidéer producerer nektar. De øvrige tilbyder forskellige andre spise- og duftstoffer. F.eks. har mindst 50 af Ecuadors Maxillaria arter et lyst pudder på læben dannet ud fra kirtelhår. Pudderet skal erstatte pollen og lokke pollensamlere til. Pudderet består af afsnørede celler, der indeholder stivelse eller olier. T.v. Maxillaria grandiflora (Foto: Mauro Rosim, Flickr). Callus på læben kan bestå af hår eller papiller, der udskiller olie. Mange insekter tygger på hårene og får derved en føde belønning for besøget i blomsten. T.h. Coelogyne cristata.
Når blomster bliver under ca. en cm i størrelse, så bliver de let overset af insekter, og derfor samles små blomster i blomsterstande, som kan ses klart på langt større afstand. Hos Oncidium truliferum tiltrækkes visse bier af blom-sterstanden, fordi de opfatter den som en bisværm.
Blomstens genkendelighed spiller også en vigtig rolle for at kunne tiltrække et bestemt insekt. Her drejer det sig om form, størrelse, farve, UV-mønster og duft. Insekter gen-kender lettere asymmetriske end symmetriske former. Især den zygomorfe eller énsymmetriske blomst, som karak-teriserer orkidéer, tiltaler insekter. Billedet viser norne, Calypso bulbosa, der er cirkumpolar i nåleskovsbæltet.
Nogle orkidéer benytter sig af mimicry for at lokke insekter til, dvs. blomsterne, ligner hunnen af et insekt, hvor hannen derfor forsøger kopulation. Flueblomst, Ophrys insectifera bestøves af gravehvepsehannerne Argogorytes mystaceus og A. fargei. Hannen foretager en pseudokopulation. Hannerne udvikles 14 dage før hunnerne og har derfor ingen at parre sig med. Blomsten imiterer på enestående vis hunhvepsens udseende, og hvad mere er, den dufter stærkere af kønshormon end hunnen. Bemærk, at de to sidestillede blosterblade i den indre krans (petaler) danner ’følehornene’. Det har vist sig, at formen ikke er særligt vigtig for at lokke hannerne til. Klippes der noget af læben, er besøgene lige hyppige, blot duftkirtlerne ikke fjernes. Der er ingen nektar i blomsten, og hanhvepsen besøger den derfor alene for at parre sig med den. Derved bliver planten afhængig af, at der samtidig er andre blomster, der kan levere føde til hvepsen. Når hunhvepsene kommer frem har flueblomstens blomstring kulmineret. Når hannen har besøgt flueblomsten dufter den som en hun, og dermed kan kopulationsmønstret blive et helt andet.
En anden form for mimicry, der også er rent snyd og be-drag, ses hos den australske orkidé Thelymitra antennifera. Den producerer ikke nektar, men den ligner en lilje, der har nektar, og derfor besøges den af nektarsøgende insekter. Foto: P Bouchard, Flickr.
Ascocenda Thai Lemon
Dendrobium lawesii
Dendrobium victoria- reginae
Coelogyne speciosa
I orkidéfamilien er alle tænkelige farver realiseret hos blomsterne. De mange farver og kombinationer af farver afspejler, at et uhyre stort antal forskellige insektarter optræder som bestøvere, men som nævnt ofte sådan, at et enkelt insekt kun bestøver en enkelt eller nogle ganske få orkidé arter. – I den forbindelse skal det nævnes, at der er betydelig forskel på, hvordan forskellige insekter opfatter farver, og deres farveopfattelse er meget forskellig fra vores. F.eks. kan mange bier ikke opfatte rødt, hvorimod visse dagsommerfugle godt kan. Generelt er insekters farvesyn forskudt i forhold til vores, så for bier er de grønne blade gråsorte, og derved kommer blomster til at fremstå lysende på mørk baggrund.
Duft er endnu en vigtig faktor for at kunne tiltrække bestøvere til orkidéer. Mange har en opfattelse af, at orkidéer ikke dufter. Det skyldes nok at Phalaenopsis hybrider er almindelige stueplanter, og de fleste dufter ikke. Herover ses t.v. den vel-duftende Zygopetalum mackayi fra Brasilien. – T.h. ses en dueurtsværmer (Deilephila elpenor) stikke snablen frem (Foto: J Mangelsdorf, Flickr). Forsøg har vist, at natsværmere rækker snablen frem i vindtunneler, når der serveres blomsterduft, og ligustersværmere (Sphinx ligustri) kan orientere sig alene efter duften uden syn. I Amazonas regnskoven i Ecuador bestøver orkidé bier (Euglossa sp.) Chondrorhyncha caudata. Han bien tiltrækkes af orkideens duftkirtler, som den samler op på bagbenene for at forføre hunnerne. Når bien forlader blomsten, sker det ofte med pollinierne fasthæftet på hovedet. – Fugle har derimod ingen lugtesans. Oppe i bjergene afløser fugle ofte insekter som bestøvere.
Nektar er et oplagt lokkemiddel. Den findes oftest i en spore på læben som hos Bakke-gøgelilje (Platanthera bifolia, t.v.) og Aerangis thompsonii (i midten), men nektarier kan også være placeret ved grunden af petaler og sepaler eller på søjlen. På fotoet af Bakke-gøgelilje kan nektar-niveauet anes i bunden af sporene. Hos A. thompsonii er læben bagud bøjet, da det er natsværmere, der bestøver den, og de står og svirrer med vingerne i luften og skal ikke bruge en landingsplads. Nektarier kan også være extraflorale og sidde på vegetative dele, som vist t.h., hvor der sidder en nektar dråbe ved grunden af hver blomsterstilk hos en Cattleya. Disse nektarier skal opfattes som en beskyttelse mod, at kravlende dyr når frem til blomsten.
Der er endnu nogle forhold, der kan lokke insekter til, men de har ingen betydning for orkidéer. Nogle arter betjener sig af reklameblomster. Det er golde blomster, der har til formål, at gøre de fertile blomster mere synlige. Det gælder f.eks. randblomsterne på kurvblomster og Hortensia (Hydrangea) vist her t.v. T.h. er det Lysichiton americanus fra Arum familien, hvor det farvede hylsterblad gør blomsterstanden synlig. Denne art har yderligere det raffinement, at kolben udvikler varme, der lokker insekterne til, her er det små fluer.
Blomstens åbningstid og dermed holdbarhed spiller også en rolle for succesfuld bestøvning. En selvbestøvet blomst behøver overhovedet ikke at åbne sig. Orkidéer udmærker sig generelt ved, at blomsterne holder meget længe. Det kan hænge sammen med, at der ofte er langt mellem de enkelte individer. Afhængigheden af et bestemt insekt bliver mindre kritisk, når blomsten er tilgængelig i lang tid. Stanhopea arter med store synlige blomster og stærk duft holder dog kun 4-5 dage. Cattleya arter holder i 5-8 uger. Phalaenopsis amabilis og hybrider med den (her Alice gloria, t.v.) kan holde i op til 4 måneder. Men det er småting mod Grammatophyllum multiflorum fra SØ-Asien, hvor holdbarheden er trekvart år. Som kontrast kan det nævnes, at flagermus bestøvede kaktus som Cereus jamacaru (t.h.) kun holder en nat.
Det koster meget i energi at holde blomsterne friske så længe, og derfor er det en fordel, hvis blomsten kan bidrage til at dække energiforbruget ved egen fotosyntese. Mange orkidéblomster er grønlige, hvilket skyldes tilstedeværelse af klorofyl. For en enkelt Cymbidium art er der også ført bevis for, at blomsten selv udfører fotosyntese. Det er givetvis også tilfældet her hos Coelogyne pandurata (Malaya-Borneo). – Både C. pandurata og Dendrobium margaritaceum (t.h.) illustrerer, at der skal være et sted, hvor insektet kan lande, når det kommer på besøg, og i en zygomorf blomst er det altid læben, der fungerer som landingsplads. Ved at sætte sig her er insektet straks rigtigt orienteret både i forhold til fødesøgning og bestøvningsprocessen. Her er både røde og gule lys som i en anden lufthavn. Hos andre arter fungerer farvede streger på læben som vejvisere. Se f.eks. de tidligere viste fotos af Calypso bulbosa, Coelogyne cristata og Zygopetalum mackayi.
Meget pollen og store støvfang er karakteristisk for vindbestøvere. Orkidéer må også producere meget pollen af hensyn til de mange frøanlæg, der skal befrugtes. Pollen overføres hos flertallet af arter på en gang klæbet sammen i pollinier, der ses herover hos Embreea rodigasiana (Columbia-Ecuador) med klæbeskiven, som har været hæftet på bestøveren. Fanget er ofte relativt stort (brunligt hos Rossioglossum grande t.h.), måske for at sikre større træfsikkerhed, når overførslen sker, så skævt placerede pollinier også kan opfanges. Pollinier og støvfangs placering er vigtig for at maximere præcisionen og samtidig minimere energiforbruget ved pollenoverførslen. Sammenvoksningen mellem det fungerende støvblad og det ene frugtblad til griffelstøtten (søjlen) tjener specielt dette formål, og den er samtidig basis for en så kraftig specialisering af blomsten, at kun en enkelt insektart evt. kun det ene køn af den pågældende insektart kan klare at udføre bestøvningen. En sådan tilpasning har krævet millioner af år at udvikle, og den er sket ved en sideløbende specialisering af plante og insekt kaldet co-evolution.
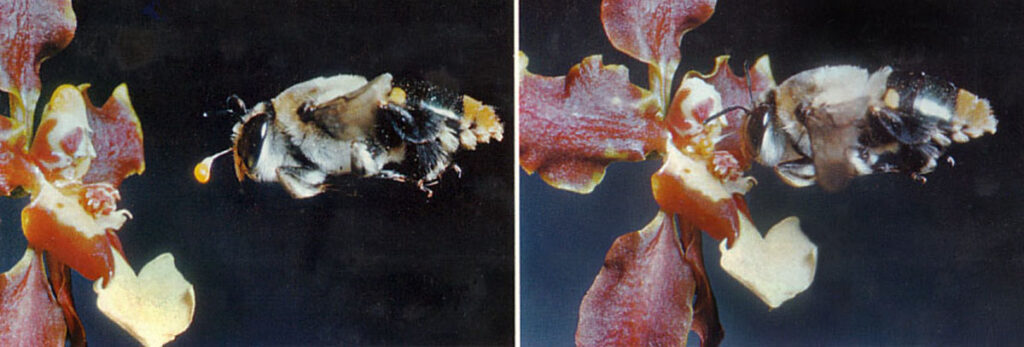
Det ses herover, at specielt læbens udformning med opadbøjede vinger begrænser bestøverens bevægelsesfrihed. Dvs, skal dyret frem til maden, så må det ind i røret, og der er kun én vej frem. Der er også kun én vej tilbage, og når bestøveren bakker ud igen, vil det modhageformede rostellum gribe fat i dyret, og polliniernes klæbeskive rives løs, så pollinierne følger med og klistres på dyret. Den viste humlebi besøger Schomburgkia superbiens i Danmark, og det er således ikke den naturlige bestøver, så det ser ud til, at pollinierne sidder uhensigtsmæssigt for langt bagud på humlebiens vinger. S. superbiens vokser som epifyt i bjergskov i Mellem-Amerika op til 2.000 m.o.h.
Samarbejdet mellem orkidéer og de bestøvende insekter går så vidt, at mange orkidéer signalerer, at nu er de blevet bestøvet, så det er ikke umagen værd at besøge blomsten, for nektaren er taget, og der produceres ikke mere. Signalerne er mangfoldige og kan bestå i farveændringer af dele af blomsten, ændringer i duften, opsvulmning af dele af blomsten, hormonproduktion m.m. – Under blomstens visning trækkes en del nedbrydningsprodukter tilbage i planten og genbruges. Visningen startes af en ethylen produktion, der sættes i gang, når støvknaphætte og pollinier fjernes med mindre blomsten er bestøvet. Undersøgelser af sådanne postbestøvnings-fænomener hos slægten Phalaenopsis førte for ca. 160 år siden til opdagelse af cellekernen og for ca. 100 år siden til opdagelsen af plantehormonet IAA, indoleddikesyre, der er et auxin. Bemærk på billedet i midten af Zygopetalum mackayi, at blomsten ikke har afleveret sine egne pollinier, men modtaget et par fra en anden blomst. Disse pollinierne er nu skrumpede, fordi pollenkornene er spiret og vokset ned gennem griffelstøtten til frøanlæggene. Samtidig er griffelstøtten svulmet op omkring støvfanget. T.h. ses den bestøvede blomst øverst, hvor frugtknuden er blevet tykkere sammenlignet med den visne ubestøvede blomst nedenunder, og petalerne er forblevet grønne, så de kan bidrage med fotosyntese.
Selvbestøvning er konstateret regelmæssigt hos ca. 3% af alle orkidé arter. Nogle er tilmed apomikter, dvs. at de kan udvikle frø uden befrugtning. I Danmark findes selvbestøvning hos flere arter af Cephalanthera. I denne slægt mangler rostellum og klæbeskive, så pollen og fang er dårligt adskilte (lim på fang). T.v. ses Hvidgul skovlilje (C. damasonium), hvor blomsterne næsten ikke åbnes. T.h. er det mygblomst (Liparis loeselii), der selvbestøves v.h.a. regndråber, der slår pollinierne ud af knappen, hvorefter de rammer den krumme læbe, så den slår pollinierne op mod støvfanget.
Man antager, at insektbestøvning er udviklet fra vindbestøvning. T.v. føres en pollensky med vinden væk fra enebær (Juniperus communis). Primitive vindbestøvede nøgenfrøede planter har ligesom vore dages nåletræer udskilt en klæbrig substans omkring frøanlæggets munding, micropylen, beregnet til at opfange de pollenkorn, der føres frem med vinden. Biller, der kravlede rundt på planterne har opdaget en næringskilde i pollen og den klæbrige væske. De har så stadig mere regelmæssigt opsøgt frøanlæggene og har ikke kunnet undgå at overføre pollenkorn fra plante til plante og derved gjort bestøvningen mere effektiv. T.h. ses unge hunkogler af Thuja occidentalis med væskedråber på nogle af frøanlæggene Foto: Plant Image Library, Flickr.
Riddergøgeurt (Orchis militaris t.v.), Stor gøgeurt (Orchis purpurea) og mange andre gøgeurter bestøves af enlige bier (modsætning til sociale bier) og humlebier.
Flueblomstens (Ophrys insectifera) samspil med gravehvepse hanner er allerede omtalt. Sydeuropæiske flueblomster som Ophrys speculum (Foto t.v.: M Freeth, Flickr) og Ophrys tenthredinifera (i midten) bruger også ligheden med insekter til at lokke hanner af forskellige bier til sig. Ophrys lutea (t.h.) bestøves af en hanbi af slægten Andrena, men her er læbens efterligning af hunnen orienteret den anden vej, dvs. hovedet vender væk fra blomstens centrum, og kopulationen foregår tilsvarende, så pollinierne afsættes på hannens bagkrop (Nederste foto: D Attenborough 1995).
Cypripedium calceolus, fruesko bestøves af 5 forskellige små arter af jordbien Andrena, der ofte ender med at falde ned i skoen. Der er gennemskinnelige partier på læbens bagvæg (rød cirkel), som viser vej til udgangen ved den røde pil, der peger på en støvknap. Fruesko og den nært beslægtede Paphiopedilum afviger fra flertallet af orkidéer ved at have to støv-knapper, én på hver side af søjlen, så bien vil ramme imod en støvknap uanset, hvilken af de to mulige udveje den vælger.
Cycnoches (Mellem- og Sydamerika) har som omtalt énkønnede blomster, der hos nogle arter er næsten ens, men hos andre meget forskelligt udformede. Her ses igen Cycnoches chlorochilon med hun t.v. (Foto: J Pestana, Flickr) og han t.h. Læben vender opefter. Euglossine er en familie af tropiske bier, hvor hannerne tiltrækkes af bestemte dufte, der har bedøvende virkning. Blomsterne indeholder sjældent nektar. Hanbien lander på læbens opadvendte side, og glider så om på undersiden, hvor den må slippe med bagbenene for at nå det berusende duftområde ved spidsen af læbens callus. Herved svinger den tunge bagkrop nedad og rammer griffelstøtten (søjlen). Det udløser pollinierne, der er under spænding. De hæftes på bien nær bagkroppens spids. Tilsvarende sker i hunblomsten, hvor pollinierne afsættes på fanget, når bien letter ved at lade sig falde fra læben. – Cycnoches egertonianum er en art med stor forskel mellem kønnene. Den besøges af hanbien Euglossa hemichlora. Læben har et hængsel, så den ydre del svinger ned under biens vægt, hvorefter bagkroppen ramler mod støvknap eller støvfang.
Stanhopea tigrina bestøves ligeledes af hanbier, der tiltrækkes af en meget kraftig duft, der produceres fra et kirtelvæv i den hule læbe. Læbens midterflig og sideflige danner en slidske, som bien rutsjer baglæns ned ad, når den er blevet godt bedøvet af duften. Passagen mellem den meget lange krumme griffelstøtte og læben er så trang, at rostellum stødes af med rygsiden af brystkassen (thorax), og pollinierne fasthæftes. Medbringes pollinier afsættes de på fanget, og da blomsten samtidig er først hanlig er selvbestøvning forhindret. Pollinierne er tykke, mens fanget er smalt og indsænket indtil det ved modenhed svulmer op.
Stanhopia arternes forskellige udformning af læbe og griffelstøtte er et godt eksempel på, at hver art benytter én eller få arter af hanbier til at udføre bestøvningen. Den tidligere viste Stanhopea cucullata har en mindre blomst end S. tigrina, men en bredere tragt mellem læbe og griffelstøtte tilpasset en større biart end den hos S. tigrina. Hvilken funktion øjepletten har, er jeg ikke klar over. Herover ses t.v. S. martiana fra Mexico og t.h. S. napoensis fra Ecuador.
Hos Gongora vender læben opad, og den er ret kompliceret udformet. Når hanbien sætter sig på læben vender bien rundt, så den kommer ned at hænge under læben, hvor den skraber efter duftstoffer. De sidder ved basis af læben, så bien skal ind mellem gaflerne, der så styrer den, når den mister fodfæstet og falder på ryggen ned på griffelstøtten, hvor pollinierne samles op på rygsiden. Billedet t.v. viser Gongora quinquenervis. Det er en ret variabel art, der findes fra Mellemamerika til Peru. Foto t.h. er af G. galeata fra Mexico. Den bestøves på samme måde, blot er hanbien en anden art.
Adskillige orkidéer udsender hunlige pheromoner for at lokke bestemte insekt hanner til. Det er allerede omtalt under bestøvningen af Flueblomst (Ophrys insectifera). Her følger nogle eksempler fra Australien. Den vingeløse hun af snyltehvepsen Neozeleboria cryptoides lægger sine æg i insekter. Når en hun er kommet frem fra sit spisekammer, er hun afhængig af at blive fodret af hannen. For at fange hans opmærksomhed kravler hun op i toppen af en plante og udsender pheromoner. Når en han finder hende parrer de sig, og han fragter derefter hunnen fra Eucalyptus træ til Eucalyptus træ, hvor hun lever af udskillelserne fra bladlus, indtil hannen afleverer hende ved et insekt, hvor hun kan lægge æg. Er orkidéen Chiloglottis trapeziformis i nærheden, går det dog galt for snyltehvepsen, for orkidéen udsender 10 gange så meget hunligt pheromon som snyltehvepse hunnen (10µg mod 1µg), og da læbens sorte callusområde imiterer en hvepsehun, vælger hannen derfor orkidéen og forsøger en psedokopulation, hvorved den samtidig sørger for at aflevere pollinier, hvis den har nogen med.
Snyltehvepsen Neozeleboria cryptoides pseudokopulerer med orkidéen Chiloglottis trapeziformis. Pilen i A peger på støvfanget. I B afleveres pollinier. Efter besøg i 4-5 blomster lærer hannen, at den tager fejl, og orkidéen må derfor hele tiden udvikle stærkere attraktive midler, så de næste generationer også kan blive bestøvet. Det er et eksempel på trinvis co-evolution. Fotos: M L de Jager & Rod Peakall 2019, Ann. Bot. 123: 347–354.
Herover ses 3 eksempler på australske hammerorkidéer. Det er orkideer, hvor læben har et hængsel, som især snyltehvepse (thynnine hvepse) aktiverer, når de sætter sig på læben. Derved smækker læbe og hveps ind mod griffelststøtten, hvor pollensække så kan afleveres eller opsamles. Det er igen pheromoner og fysisk lighed med en hun, der tiltrækker han hvepsen. Øverst t.v. ses Drakaea thynniphila (Foto: P Bouchard, Flickr). Nederst t.v. er det Drakea livida (Foto: Jean & Fred, Flickr). De øvrige fire fotos er Drakaea glyptodon (Foto: B Bohman et al. 2014, New Phytol 203,3). Den bestøves af den hanlige thynnine hveps, Zaspilothynnus trilobatus. (a) viser blomsten med den leddelte læbe, hvor den yderste del imiterer hun hvepsen. (b) En han forsøger at kopulere med en attrap oversmurt med en syntetisk blanding af pyraziner. (c) En hvepse han forsøger at flyve væk med den falske hun, hvilket medfører at hængslet aktiveres, så hvepsen slynges over mod griffelstøtten. (d) Processen er fuldført.
Pterostylis banksii fra New Zealand bestøves af enlige hanner af bier og humlebier. De op til 5 cm store blomster har også en bevægelig læbe (6). 1-3 er sepaler, 4-5 petaler og 7 er frugtknuden. Læben har et led ved grunden, og når et insekt sætter sig på læben, vil dets vægt få læben til at svinge ind i blomsten, hvor insektet, oftest en flue, holdes fanget i ½-2½ timer, før læben atter giver efter, og når insektet så kravler ud, vil det på vejen opfange og/eller aflevere pollinier. Foto: B Spragg, Flickr.
Anguloa clowesii fra Columbia og Venezuela er en anden orkidé med bevægelig læbe. De to fotos viser læben i yderpositionerne.
Mange andre orkidéer bedøver bestøveren. Den syd- og mellemamerikanske slægt Coryanthes er nok den mest komplicerede orkidé blomst, hvor bestøvningen bygger på bedøvelse af en hanbi. To kirtler ved griffelstøttens basis drypper væske (rød cirkel) ned i den skålformede læbe. Hannen sætter sig på den nærmest grotesk store blomst (over 20 cm) ved læbens opsvulmede basis tiltrukket af nektar duft. Der er ingen nektar, men bien parfumerer sig så med et hormonlignende stof, der produceres på læbens øverste (basale) del. Herved bliver hanbien attraktiv for hunnerne. Duften bedøver imidlertid hanbien, og den dratter ned i den væskefyldte læbe, hvor den må kæmpe sig ud for ikke at drukne. Den kan ikke kravle op ad læbens vægge, da de er glatte som indersiden af en Sarracenia kande. Der er selvfølgelig kun én vej ud forbi spidsen af griffelstøtten med pollinierne.
Passagen forbi griffelstøtten er så snæver, at det er hårdt arbejde for bien af mase sig igennem, men det sikrer til gengæld, at pollinierne hæftes fast eller tilsvarende afsættes på fanget, hvis dyret har nogen med sig. Fanget passeres først.
Det er tidligere nævnt, at små blomster samles i blomsterstande for at gøre dem mere synlige, og mange af dem bestøves af sociale bier. Det gælder som nævnt f.eks. Oncidium truliferum, hvor blomsterstanden kan opfattes som en bisværm. Det er også sociale bier, der bestøver vores hjemlige gøgeurter som Plettet gøgeurt (Dactylorhiza maculata, t.v.), Salep-gøgeurt (Orchis morio, midten) og Tyndakset gøgeurt (Orchis mascula, t.h.). De to første bestøves især af honningbien Apis mellifera.
Skov-hullæbe (Epipactis helleborine) bestøves af skovgedehams (Vespa sylvestris). Nektaren delvist bedøver hvepsen, så den efter besøget har svært ved at flyve. Den tumler derfor ofte bare videre nedad til næste blomst på skaftet. T.h. ses den falde ud af den nederste blomst med pollinier på panden. Skovhullæbe har lang blomstringstid i juli-august samt atypisk vedvarende nektar produktion. Sump-hullæbe (Epipactis palustris) besøges også af forskellige hvepse inkl. gedehams og bier. Foto af hvepse: H Tybjerg.
Hos Oncidium planilabre (herover) og O. hyphaematicum går droner til angreb, idet de opfatter blomsten som et fjendtligt insekt. Bien sætter sig ikke på blomsten, men buser hovedkulds ind i den for at jage den væk. Foto: Pijl & Dodson 1966.
Ægbladet fliglæbe (Listera ovata) bruger snyltehvepse og tæger som bestøvere. Blomsten har nektar i en åben grube på læben. Når insektet rammer mod rostellum (griffelstøttens næb) eksploderer det, og der frigøres klæbestof, der hæfter pollinierne fast på insektet. Det reagerer med forskrækkelse og forlader straks blomsten, men blot for at sætte sig på en anden, så chancen for fremmedbestøvning er kraftigt forøget. Fluer kan også bestøve Listera.
Nogle orkidéer bestøves af dagsommerfugle. Herhjemme gælder det horndrager (Anacamptis pyramidalis, t.v.). Blomsten har en relativt lang og meget snæver spore som dagsommerfugle kan nå ned i. Den røde farve er også i orden, idet mange dagsommerfugle kan opfatte rødt. Blomsterstanden er så tætblomstret, at den som helhed kan fungere som en landingsplads. Sommerfuglen sætter sig på toppen og drejer rundt om sig selv for at nå blomsterne med snablen ligesom kejserkåben (Argynnis paphia) gør det på Ager-tidsel (Circium arvense, foto t.h.). – Bakke-gøgeurt (Orchis ustulata, midten) siges også at være mest besøgt af dagsommerfugle. Indgangen til blomsten er for snæver til bier. På Öland er den set bestøvet af store fluer, hvor pollinierne sætter sig på snablen.
Psycopsis papilio kaldet sommerfugle orkidé bestøves af dagsommerfugle. Der er fire arter i slægten udbredt fra Caribien til Peru. Blomsterne kommer successivt hævet langt over bladene på et op til 70 cm langt skaft, hvor de vibrerer i vinden og efterligner derved en flyvende hun sommerfugl. Det tiltrækker han sommerfugle, der parrer sig med blomsten og samtidig bestøver den. Blomstens sommerfugle mimicry har yderligere den effekt at tiltrække parasittiske insekter, der er på jagt efter sommerfugle, de kan lægge æg i.
Bakke-gøgelilje (Platanthera bifolia, t.v. og i midten øverst), er en typisk natsværmer blomst. Den er hvid, den dufter sødt om aftenen, og den har en lang spore med rigelig nektar. Da pollinierne sidder meget tæt sammen lige ved sporens indgang, får natsværmeren, evt. en ugle, pollinierne på snablen. Hos Skov-gøgelilje (Platanthera chlorantha, i midten nederst) får natsværmeren pollinierne på øjnene, fordi pollinierne sidder langt fra hinanden, og da fanget hos Bakke-gøgelilje er ganske lille og placeret lige ved sporemundingen, så det kun kan rammes af pollinier siddende på snablen, så vil de to arter kun sjældent hybridisere, selv om de jævnligt besøges af samme insekt. Hos Bakke-gøgeliljen bøjer polliniets stilk tilmed fremover, så pollinierne blot føres med ind og ud af sporemundingen hos Skov-gøgelilje. Læg også mærke til læben. Den er slået ned og fungerer tydeligvis ikke som nogen landingsplads, idet natsværmere ikke lander, men står og svirrer foran blomsten. – Habenaria dilatata (t.h.), fra Nordamerika er også natsværmer bestøvet.
Aerangis thompsonii t.v. med den lange spore og tilbageslået læbe er en typisk natsværmer bestøvet blomst. Det samme gælder Angraecum eburneum, hvor blomsten atypisk er drejet, så læben heller ikke fungerer som landingsplads, idet den vender opad. Bestøveren står og svirrer foran blomsten, og snablen styres ind i den forholdsvis korte spore ved hjælp af renden i griffelstøtten. Begge slægter er afrikanske.
Angraecum sesquipedale fra Madagascar er en bemærkelsesværdig natsværmer bestøvet orkidé med en lang spore på omkring 30 cm. Arten blev beskrevet af Darwin i 1862, og han forudsagde ud fra sine teorier om tilpasning og co-evolution mellem arterne, at der måtte eksistere en natsværmer med en snabel af samme længde, der kunne bestøve planten. 40 år senere opdagede man så det rette dyr Xanthopan morganii f. praedicta. Det var dog først i 1997, at man observerede, at orkideen blev bestøvet som forudsagt. Xanthopan foto: kqedquest, Flickr.
Nogle orkidéer er tilpasset flue bestøvning. Der er to hovedformer for fluebestøvning. Ved den simpleste form har fluen fri adgang både til og fra blomsten. Det gælder f.eks. Bulbophyllum, den mest udbredte slægt med over 2000 arter. Herover B. lemniscatoides hvor hvert blosterblad er trukket ud i et langt smalt vedhæng, der duver i vinden, hvilket er attraktivt for visse fluer. Andre arter lugter desuden råddent af forskellige emner, der hver især tiltrækker en enkelt flue art. – I Amerika er slægten Pleurothallis (t.h.) overvejende fluebestøvet.
Den mere avancerede form for flue-bestøvning er repræsenteret hos Paphiopedilum, der har bygningstræk, der minder om andre fluefælder (tragt med tag over som hos Sarracenia, stribet tag som hos Arisaema (Araceae)). Grøn, brun, mørkerød, hvid og purpur/violet er de dominerende farver. Dertil kommer hos nogle en dårlig lugt. Fluer, biller og hos nogle arter bier kommer ind af den store åbning i læben og forlader fælden ad de små åbninger ved hver af de to støvdragere, som tidligere beskrevet for vores hjemlige fruesko (Cypripedium calceolus), der især besøges af små jordbier. Her er t.v. vist Paphiopedilum maudiae var. coloratum, og t.h. en hybrid Paphiopedilum ’Lynleigh Koopowitz’.
Pletter og behåring som hos Paphiopedilum hirsutissimum (t.v.) fra Indien-Thailand og Cambodia samt P. sukhakulii (t.h.) fra NØ-Thailand er også tillokkende for fluer.
Rhizanthella gardneri bestøves af termitter. Orkidéen blev fundet første gang i 1928 af en landmand i SV-Australien, da der pludselig vældede udsprungne blomster op af jorden, mens han pløjede sin mark. Planten vokser og blomstrer fuldstændig under jorden. Den har intet chlorofyl og får sin næring gennem svampe, der har mycorrhiza med grønne autotrofe planter. Orkidéen er således en mycoheterotrof plante ligesom rederod (Neottia nidus-avis) og snylterod (Monotropa hypopitys) i Danmark. De ca. 150 tæt pakkede små blomster tiltrækker termitter som bestøvere, og de dufter så kraftigt, at mennesket kan opfange duften gennem jorden. Man kender kun 6 populationer, så arten betragtes i stor fare for at uddø. Foto: Jean & Fred, Flickr.
Den eneste dyregruppe ud over insekterne, der spiller en rolle for bestøvning af orkidéer er fugle, specielt de amerikanske kolibrier, der kaldes hummingbirds. Der er ingen duft i fugleblomster, men stærke røde, orange eller gule farver, og der er masser af nektar i solide sporer eller i nektarier indbygget i frugtvæggen. Der er ikke brug for en landingsplads, så læben er oftest smal og rørformet som hos den mellemamerikanske Prosthechea vitellina (herover) tidligere kendt under navnene Epidendrum vitellinum og Encyclia vitellina (Foto: H Tybjerg). Interessant nok findes der også en hvid form i naturen, men om den så trods manglende duft er natsværmer bestøvet ved jeg ikke. Kolibri foto: K Bondy, Flickr.
Cattleya aurantiaca (t.v.) fra Mellem-Amerika med den smalle rørformede orangegule læbe rapporteres bestøvet af kolibrier, mens de fleste andre Cattleya blomster bestøves af bier. – Sophronitis coccinia (= Cattleya grandiflora, t.h.) fra Brasilien anses også for at være bestøvet af kolibrier.
Cattleya aclandiae, der gror i Bahia provinsen i Brasilien, kombinerer karakterer for både fugle- og insektbestøvning, idet læben fungerer som landingsplads, og der er duft i blomsten samtidig med, at søjlen danner et rødt rør, der passer til tungen på en kolibri.
Der er som nævnt mindst 25.000 orkidé arter, og dertil kommer en lang række naturlige hybrider, så denne gennemgang af orkideernes bestøvningsbiologi beskriver kun en begrænset del af den variation, der forekommer. Det kan tilføjes, at der er mindst 20 millioner insekter at tage af, hvoraf mindre end 950.00 er navngivet. Der kendes ca. 9.000 fuglearter, men forsvinder blot én art af bestøverne, kan det let medføre, at en orkidé art også går tabt.