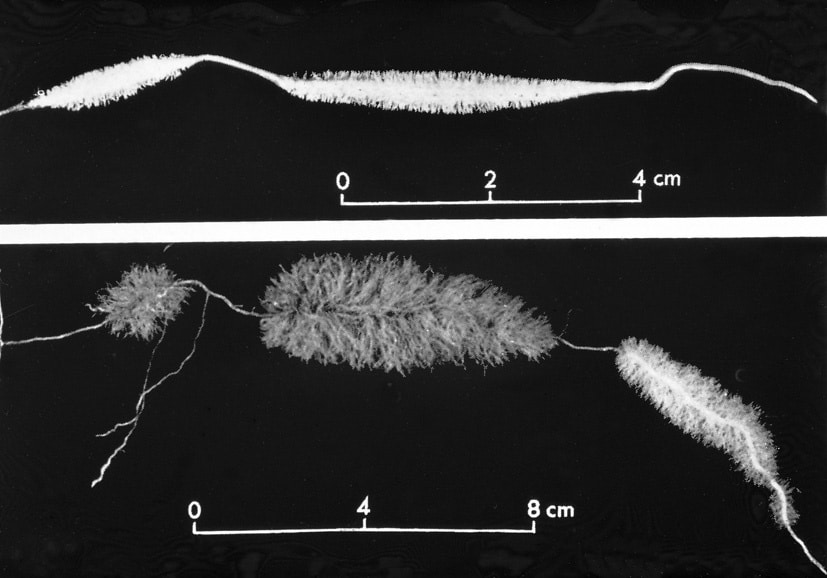
Rundbladet soldug er den mest udbredte soldug art i Danmark. Den er cirkumpolar inklusive det sydlige Grønland. Den trives fint med konkurrence fra tørvemos (Sphagnum). På billedet th. vokser soldug tæt rundt om en grundvands regulere klitlavning i den rødbrune bræmme, hvor der hverken er for vådt eller tørt.
Drosera rotundifolia blomstrer i august. De 5-tallige blomster er kun åbne midt på dagen, og de sidder højt hævet over fælderne. I tørt og solrigt vejr kan slimen på tentaklerne tørre ud. På billedet herunder ses i midten et blad foldet sammen om et bytte som resultat af den på hovedsiden omtalte auxin regulerede vækstbevægelse.
Langbladet soldug - Drosera anglica
Langbladet soldug har i Danmark kun få voksesteder på højmoser og i fugtige klitheder i Jylland. Dens totale udbredelse er næsten cirkumpolar. Den findes dog ikke i Island og Grønland, men i Nordamerika når den op i Alaska. Der findes også nogle subtropiske forekomster på Hawaii og i det sydlige Europa og sydlige Japan. Den trives fint i sure omgivelser mellem Sphagnum. Den overvintrer ved vinterknopper kaldet hibernakler. Arten er selvbestøende og menes opstået som en diploid hybrid mellem den nordamerikanske D. linearis og D. rotundifolia. Drosera anglica x obovata er en steril hybrid med D. rotundifolia. Hybriden forveksles undertiden med Drosera intermedia.
Liden soldug - Drosera intermedia
Liden soldug findes især i moser, hvor den står under vådere forhold end de to øvrige danske arter, rundbladet og langbladet soldug. Den har hovedudbredelse i det vestlige Europa og østlige Nordamerika, men når ned i det nordlige Sydamerika. I tempereret klima overvintrer den ved hibernakler. Bladene er oprette, og formen er intermediær mellem de to førnævnte arter. Blomsterstanden virker sidestillet, idet den er bøjet til siden, mens den står opret centralt i rosetten hos de to andre danske arter. Bomstrer fra juni til august. På billedet t.h. vokser den sammen med blærerod Utricularia australis. – Drosera anglica x obovata er en steril hybrid med D. rotundifolia. Hybriden forveksles undertiden med Drosera intermedia.
Xerofyt kommer af græsk og betyder en tørketilpasset plante. En xerofyt kan vokse, hvor nedbøren er lav og jorden hurtigt tørrer ud samtidig med, at der ofte er varmt, dvs. på heder, i klitter, halv- og helørkner. Xerofytter klarer sig under disse forhold fordi, de har en række anatomiske, morfologiske og fysiologiske tilpasninger, der nedsætter deres transpiration og dermed vandforbruget, og desuden er mange i stand til at opmagasinere vand i rødder, stængler eller blade, som der kan tæres på i tørkeperioder.
I det følgende gennemgås de forskellige tilpasninger med det lille stedsegrønne australske træ Hakea drupacea som eksempel, og derefter følger eksempler fra en række andre planter fra både indland og udland. H. drupacea hed oprindeligt H. suaveolens, og jeg har skrevet flere afhandlinger om den under det navn. Det er ikke en udpræget xerofyt, idet den vokser under relativt gode nedbørsforhold i et kystnært bælte langs det sydvestlige og sydlige Australien, men dog på sandet veldrænet jord, og den skal klare en tørtid. Den er introduceret flere steder i verden, bl.a. i Sydafrika og på New Zealand. På billedet t.v. blomstrer den i et drivhus i Københavns Botaniske Have.
Når Hakea drupacea alligevel besidder en lang række tørketilpassede (xeromorfe) karakterer, forklares det ved, at dens forfædre har vokset under forhold, hvor der har været brug for karaktererne, og da de på ingen måde hæmmer arten under de nuværende forhold, så selekteres der ikke imod karaktererne.
Rodsystemet hos Hakea absorberer vand fra jorden særligt effektivt, idet der er udviklet såkaldte proteoide rødder. Det er afsnit af rødderne, hvor der anlægges et stort antal tætsiddende korte siderødder, der giver en kraftig forøgelse af røddernes absorberende overflade som vist på figuren t.h. (Lamont 1972).
Generelt har xerofytter et dybt og vidt udbredt rodsystem, men det kan også være overfladenært og indrettet til at optage regn, der kun når nogle få cm ned i jorden. F.eks. når rødderne hos en blot 25 cm høj Opuntia kaktus mere end 125 cm ud til siderne. De vandledende kars diameter i lange rødder er ofte ekstra stor for at nedsætte diffusions modstanden.
Det nåleformede cylindriske blad/bladafsnit er en xeromorf karakter, som mange planter benytter sig af, når der er brug for at spare på vandet og minimere fordampningen fra bladet, dvs. mindske transpirationen. T.v. ses Hakea drupacea sammen med en sydafrikansk xerofyt, Hymenolepis parvi-flora, der har samme xeromorfe bladform. De hjemlige nåletræer er et andet oplagt eksempel. De lider normalt ikke af vandmangel i vækstsæsonen, men når jorden er frosset om vinteren, har de samme problemer med vandforsyningen som xerofytterne.
Herover ses bladfølgen i anlægsrækkefølge hos Hakea drupacea fra kimblad til den endelige bladform med cylindriske nåleformede bladafsnit. Målestokken t.v. er 2 cm. Hakea spirer på et tidspunkt, hvor jorden ikke er vandlidende, og de små planter vil ofte være omgivet af anden beskyttende vegetation, så det er først, når planten bliver højere og kommer op i mere tør luft samtidig med, at solen opvarmer bladene, at der er behov for den xeromorfe bladform.
Transpirationen stiger med bladets opvarmning. Derfor er mikrophylli med små smalle blade en fordel for xerofytter. De to figurer herover illustrerer, hvor meget højere bladtemperaturen kan blive i forhold til den omgivende luft langs de røde linier på blade med lukkede spalteåbninger. Hvis de røde områder længst væk fra ledningsstrengene reduceres væk på det brede blad, som det er sket hos Hakea, opnås en mindre opvarmning af bladet (punkteret kurve). Kurvens form hænger sammen med tilstedeværelsen at et grænselag, hvor energiudvekslingen med omgivelser går hurtigst, hvor grænselaget er tyndest, dvs. langs bladranden. Det er årsagen til, at rimfrost først sætter sig på bladranden, og hår, der rager ud gennem grænselaget.
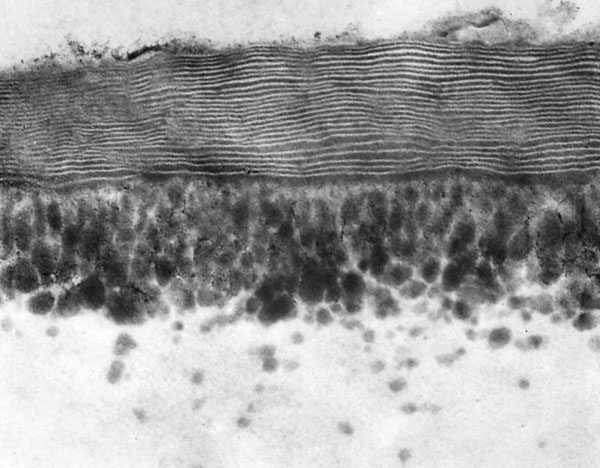
Transpiration fra blade foregår dels gennem epidermiscellernes ydervæg og dels gennem spalteåbninger, hvis åbningsgrad kan reguleres. Transpiration gennem ydervæggen er derimod en fast størrelse, der afhænger af ydervæggens konstruktion. Det yderste lag i denne væg kaldes kutikula, og laget er hos xerofytter næsten altid af en lamellær type, hvor lag af cutin og lag af voks veksler mellem hinanden. Det er begge fedtstoffer, og hos Hakea drupacea er den lamellære kutikula ekstraordinær tyk. Billedet t.h. er fra et blad i vækst. Der er konstateret op til 44 lameller i udvoksede blade. De mørke plamager er cutiner på vej til at forøge tykkelsen af den lamellære kutikula indefra. Hos andre planter er kutikula uden eller med få lameller og derfor ikke så diffusionstæt.
Skemaet t.v. viser de modstande, der skal overvindes under vandets transport gennem planten, til det fordamper fra bladene gennem spalteåbninger (stomata) og kutikula. Nogle er faste modstande som Rc (kutikula) andre variable som Rs (stomata). De ikke variable modstande er i Fick’s diffusionslov sat til K. Herefter viser ligningen, at den kontrol stomata får over transpirationsraten (T) afhænger af størrelsen på grænselagets modstand Rg. I vind med et tyndt grænselag får stomatas åbningsgrad størst indflydelse på transpirationens størrelse. Δψ er koncentrationsgradienten.
Bladenes spalteåbninger (stomata) tjener til udveksling af kuldioxyd, ilt og vanddamp med omgivelserne. De består af to specialiserede epidermisceller kaldet læbeceller, der kan ændre form styret af trykændringer, hvorved spalten kan åbne eller lukke. Stomata sidder oftest i niveau med de omgivende epidermisceller, men hos Hakea og andre xerofytter er læbecellerne indsænkede. Derved forlænges diffusionsvejen for vandmolekylerne, før de er uden for grænselaget, og det nedsætter transpirationsraten. Diagrammet viser fordelingen af tykke og tynde eftergivende vægge, der strækkes, når trykket stiger i læbecellen, så spalten åbnes. De to fotos af H. drupacea viser, hvor dybt indsænkede læbecellerne er. Det håndskårne snit herover er farvet rødt for fedtstoffer og illustrerer, hvor omfattende cutiniseringen af epidermis ydervæggene er.
Hakea drupacea og formentlig alle arter i slægten er i stand til at sætte stomata permanent ud af funktion og derved reducere transpirationen. Det sker efterhånden som bladene bliver ældre og i takt med, at fotosynteseapparatet bliver mindre effektivt og derfor kan få tilstrækkeligt CO2 gennem færre stomata. I takt med bladets alder sker der en redifferentiering af nogle palisadeceller omkring en spalteåbning. De ellers færdigt udvoksede celler, hvis primære funktion er fotosyntese, begynder at vokse og dele sig én til fire gange, så de efterhånden udfylder det substomatære hulrum og til sidst blokerer spalten mellem læbecellerne permanent. De redifferentierede palisadeceller kaldes tylosoider. Min undersøgelse viste, at i 4. bladgeneration var 6% af stomata blokerede, i 6. generation 12% og i 7. generation op til 50%. Bladene tabes i 8. generation. Den rødbrune farvereaktion i cellerne t.h. skyldes garvestof oplagret i den centrale vakuole. Blokering af ældre stomata sker hos mange andre xerofyter, men det sker ved andre metoder som blokering af stomata fra ydersiden med voks, lukning af spalten ved tab af læbecellernes evne til formændring eller ved korkdannelse.
Meget styrkevæv er en vigtig xeromorf karakter, og Hakea drupacea er velforsynet. Styrkevæv består af tykvæggede celler med forveddede vægge, dvs der er indlejret lignin mellem cellulosefibrillerne. Styrkevævs-celler kan være langstrakte og kaldes da taver. På blad tværsnittet t.v. lyser de forveddede cellevægge op i fluorescens mikroskopet, og taverne ses som mere eller mindre runde celler placeret omkring ledningsstrengene. De vandførende celler (kar og tracheider) i ledningsstrengene har også forveddede cellevægge, så cellerne får derved en dobbelt funktion.
Andre styrkevævsceller kaldet sclereider er stærkt forgrenede. Hos Hakea er de søjleformede med forgrenede ender. De er placeret mellem palisadecellerne. Styrkevævet skal forhindre, at bladet klapper sammen under vandstres, for sker det, går både fotosyntesen og transporten af vand og næringsstoffer i ledningsvævet mere eller mindre i stå.
Hakea’s tykvæggede sclereider er kun moderat ligni-ficerede, og i forgreningerne kun svagt lignificerede, så der er stadig hulrum i cellevæggen, hvor der kan transporteres vand. De to fotos herover fra et lys- og et fasekontrast mikroskop (t.h.) viser, at slereidernes forgreninger ligger klinet op mod ledningsvævets tracheider (med ringformede vægfortykkelser). Sclereiderne får derved også en funktion som ledningsstrengenes forlængede arme, og deres betydning for forsyning af palisadecellerne med vand tydeliggøres af forsøget t.v., hvor en kvist har opsuget det fluorescerende stof berberin sulfat.
Hos mange xerofytter er bladene dækket af tætsiddende luftfyldte døde hår. De reflekterer stråling og reducerer dermed bladtemperaturen, og de øger grænselagets tykkelse og reducerer dermed transpirationen. Hos Hakea drupacea er unge ikke udvoksede blade dækket af T-formede hår, men de er levende med en ekstremt tynd kutikula, så de øger bladets transpiration. Det synes i modstrid med at være xerofyt, men for Hakea gælder det om, at de unge voksende blade hurtigst muligt når det udvoksede stadium. T-hårede anlægges før spalteåbningerne er færdige og i stand til at drive transpirationsstrømmen. Det sørger T-hårene så for, og det sikrer, at der føres tilstrækkeligt med vand og næringsstoffer frem til det voksende blad.
Når stomata bliver funktionsdygtige og i stand til at styre transpirationen, bliver T-hårene skadelige, og de bortskaffes ved, at den yderste celle afkastes, så kun stilkcellen bliver tilbage. Fig. A viser det unge levende hår, hvor stilkcellens ydervæg er kutiniseret (sort). Fig. B viser ydercellen er kollapset, fordi stilkcellen på hele indersiden har dannet en kork-væg (grå), og dermed lukkes også porefelterne (C) mellem stilk og ydercellen, så ydercellen dør og løsnes fra stilkcellen. D viser, hvor tæt hårene dækker de voksende blade. E viser to døde stilkceller. F viser overfladen på et T-hår.
Øverst t.h. har et ungt blad fået en dråbe neutralrødt på overfladen, og farvestoffet er optaget i T-hårene, men der kommer ikke noget ind i bladet, så det kan afvises, at hårene skal føre dug eller regn ind i bladet. T.h. står et skud med blade i vækst (lysegrønne) og absorberer berberinsulfat. Efter et par timer er det fluorescerende stof ført med vandet ud i T-hårene, som en indikation for, at de levende hår driver vandstrømmen i det unge voksende blad.
Der er endnu et par xeromorfe karakterer hos Hakea drupacea. Det drejer sig om vandvæv, kompakt bladkød (mesofyl) og garvestoffer. T.h. vises et diagram af et såkaldt mesomorft blad hos eg (Quercus), der normalt ikke har problemer med vandforsyningen i vækstperioden. Bladkødet mellem de to epidermer består her af to lag palisadestillede celler med grønkorn og et svampvæv med mindre celler også med grønkorn, men med større luftfyldte intercellulærrum mellem sig. Hos Hakea er der to lag ens lange palisadeceller med grønkorn, men ikke noget svampvæv. Hovedparten af den centrale del af det cylindriske blad består af mere eller mindre isodiametriske celler uden grønkorn, men med store centrale vakuoler, der primært opmagasinerer vand, som palisadecellerne kan trække på, når planten er under vandstres. Der er ingen intercellulærrum i vandvævet, da stomata er placeret tættest muligt på palisadecellerne, og CO2 skal ikke transporteres tværs gennem bladet som hos eg. Palisadecellerne er slanke, så endevæggen har et lille areal for sollysets indtrængen i cellen modsat skyggeplanters palisadeceller, der er tragtformede med en bred endevæg. Xerofytter gror ofte under forhold, hvor lysintensiteten er så høj, at grønkornene skal skærmes for ikke at blive ødelagt af overbelysning. Det hjælper garvestofferne (tanniner) med til. Produktionen af garvestof fortsætter gennem bladets levetid. Først fyldes det ydre lag palisadeceller, og i 2-3. vækstsæson oplagres tanniner også i det indre lag palisadeceller. Til sidst kan det nævnes, at palisadeceller der støder op til de substomatære hulrum er beklædt med en meget tynd kutikula (300-400Å, 1Å = 10-4 µm).
Mammillaria plumosa
Oreocereus sp.
Epostoa lanata
Mange xerofytter er beklædt med døde luftfyldte hår. Det har tre positive betydninger for planten. En tæt behåring øger grænselagets tykkelse og nedsætter dermed transpirationen, og behåringen tilbagekaster en del af solstrålingen, så bladet eller stænglen opvarmes mindre, og endelig skærmes grønkornene mod overbelysning. Bemærk hos Epostoa lanata, at det især er skudspidsens unge ikke udvoksede celler, der beskyttes. De viste kaktusarter er samtidig et eksempel på sukkulens som xeromorf karakter. Sukkulente plantedele er fortykkede grundet tilstedeværelsen af et pladskrævende vandvæv. Både rødder, stængler og blade kan være sukkulente og opmagasinere vand, som assimilationsvævet med grønkornene kan tære på i tørkeperioder.
Hårenes udformning kan være meget forskellig fra xerofyt til xerofyt. Hos kaktus er de som regel lange én- eller flercellede hår. Hos havtorn (Hippophae rhamnoides) er de skjoldformede med en stilk og et mangecellet hovede, der overlapper nabohårene, så der skabes et rum med høj luftfugtighed under hårlaget og en lang vej for vandmolekylernes diffusion ud i det frie luftrum.
Rhododendron har også skjoldformede hår på bladets underside, hvor stomata er placeret, men skjoldhårene når ikke ind over hinanden og danner et ‘lukket’ rum som hos havtorn. Alligevel nedsætter de effektivt den stomatære transpiration, idet stomata er samlet i en grube under skjoldet, mens der ikke er spalteåbninger i områderne mellem skjoldhårene. Det hjælper også, at bladene ruller sig sammen under tørke.
Hos hjælme (Ammophila arenaria) er bladene kun flade i regnvejr og ved høj luftfugtighed, ellers er de indrullede, som det kaldes, når det er bladets overside, der lukkes inde. Hjælme har kun spalteåbninger på oversiden. I bunden af furerne mellem bladribberne på oversiden sidder der nogle såkaldte ‘bulliform cells’, hvor trykket kan reguleres ved ind- og udpumpning af vand, så bladpladen åbnes eller lukkes. Bemærk også det veludviklede vandvæv.
Lyngfamilien (Ericaceae) og revling-familien (Empetraceae) gemmer også stomata i huler fremkommet ved tilbagerulning eller asymmetrisk vækst af bladets over- og underside. Indgangen til hulen er ofte delvist blokeret af hår. Øverst t.v. er det revling (Empetrum nigrum), derefter kantlyng (Cassiope tetragona), der ses blomstrende t.h., og nederst hedelyng (Calluna vulgaris). Bladene repræsenterer også microphylli samt næste emne, kirtelhår der udskiller æteriske olier.
Hymenolepis parviflora (tidligere Athanasia parviflora) blev nævnt i starten af artiklen som en sydafrikansk art med samme bladmorfologi som Hakea drupacea. Bladene hos Hymenolepis har som yngre et tæt lag af grenede dækhår, der fungerer som beskrevet for kaktus. T.h. er nogle af dækhårene fjernet på et udvokset blad, så man kan se spalteåbningerne. Hymenolepis har desuden sekrethuler (3 ses på billedet t.v.) og kirtelhår, der udskiller æteriske olier.
Æteriske oliers effekt på transpirationen kan illustreres ved det simple forsøg vist herover t.v. Æteriske olier reducerer vands overfladespænding og trænger derved vandfasen længere tilbage i kapillærrøret og tilsvarende i hulrummene mellem cellevæggens cellulosefibriller, dvs. transpirationsmodstanden øges. De to fotos viser, at kutikula løsnes og spændes ud som en boble (asteriks) under sekretionen af de æteriske olier både i sekrethulerne og på kirtelhårene hos Hymenolepis. Det ender med boblerne sprænges.
Ovenstående figur viser udviklingen af de to typer hår hos Hymenolepis parviflora fra en fælles modercelle i epidermis. Øverst ses kirtelhårets og nederst det grenede dækhårs udvikling. I begge tilfælde ender det med, at hårenes stilkceller er dybt indsænkede i det udvoksede blad, og i begge tilfælde dør cellerne i hårets hoved. Hos kirtelhåret cutiniseres og forkorker (suberiniseres) den yderste stilkcelle, så der lukkes af til hovedets celler, der dør, når de æteriske olier er udskilt. Den nedre stilkcelle synes også at have mistet betydning, men hos det grenede dækhår sker der en bemærkelsesværdig udvikling vist nedenfor.
Herover vises de indsænkede grenede dækhår på et udvokset blad af Hymenolepis parviflora, dels set i lysmikroskopet og dels i transmissions elektronmikroskopet. Øverst t.h. ses et tværsnit af den næstyderste stilkcelle. Farvefotoet t.v. er et fladesnit af et udvokset blad, så palisadeceller og hårtragte ses i tværsnit. Der er ca. 300 tragte per mm2, heraf ca. 30 med kirtelhår. I de fleste tragte findes kun et hår, men enkelte rummer to hår, og de kan være af forskellig type. Nederste foto viser et udpræpareret dækhår med døde kollapsede grenceller og to levende stilkceller.
Det er bemærkelsesværdigt, at de grenede dækhårs stilkceller er udstyret med et stort antal mitochondrier og en særdeles veludviklet labyrintvæg, der forøger cellemembranens areal mere end 20 gange, men samtidig har cellevæggen en relativt tyk kutikula. En forøgelse af cellemembranen betyder normalt, at der foregår en betydelig aktiv transport på tværs af membranen, enten en sekretion eller en absorption. Desværre fik jeg ikke mulighed for at fuldføre undersøgelsen, men da grencellerne falder af på det udvoksede blad, er det tænkeligt, at behåringen har samme funktion som hos Hakea drupacea, dvs. at sikre en tilstrækkelig transpirationstrøm under bladenes vækst samt beskytte mod overbelysning. Om stilkcellerne kan ende med at være vandabsorberende er uvist.
Cakile maritima, strandkål
Senecio mandraliscae
Eucalyptus cloeziana
Mange xerofytter har uden på kutikula et lag voks kaldet epikutikulært voks, der ofte ses som et blåligt skær på bladoverfladen. På Senecio herover er vokset strøget af med en finger på et enkelt blad og chlorofyllets grønne farve træder derfor frem. Funktionen af det epikutikulære voks er at tilbagekaste en del af solstrålingen og dermed reducere bladets opvarmning og mindske faren for overbelysning. Den farve, det epikutikulære voks giver planten set med det menneskelige øje, afhænger af voksstubbenes form og orienteringen af vokskrystallerne i stubbene som antydet på Eucalyptus figuren fra Matin og Juniper 1970.
Som eksempel på vandabsorberende hår kan Bromeliaceerne fremhæves. Her er bl.a. Tillandsia usneoides en epifytisk xerofyt uden rødder, så den skal have al næring gennem bladenes absorberende skjoldhår. T.v. ses planten i tør og våd tilstand. I midten de tæt pakkede skjoldhår, og t.h. er hovedets celler døde og luftfyldte, men når det regner, suges vand og næring op ved kapillær kræfter og føres derefter ind i bladet af de levende stilkceller. Under vandstress dækker skjoldet over stomata, øger diffusionsmodstanden i grænselaget og reflekterer solstråling.
Dasylirion sp.
Psila sp.
Bredemeyera colletioides
Indsænkede stomata er allerede nævnt som en xeromorf karakter. Her vises et par eksempler mere. Dasylirion er nordamerikanske Yucca-lignende ørkenplanter, mens de to andre er sydamerikanske steppeplanter med microphylle blade uden særlig betydning for fotosyntesen, der er overtaget af stænglerne, hvor den primære bark (cortex) består af palisadeceller med grønkorn. Psila er dækket af epikutikulært voks, og den har ligesom Bredemayera dybt indsænkede stomata. Begge slægter er sydamerikanske. De sort-hvide fotos er fra studier af T Böcher og O Lyshede 1971.
Alluaudia procera
Euphorbia fimbriata
Cereus jamacaru
Bladstillingen kan have betydning for at reducere transpirationen. Det gælder f.eks. for Alluaudia fra Madagascar. Her er de små blade kantstillede for at reducere indstrålingen og dermed bladenes opvarmning. I den lange tørtid smides bladene, og Alluaudia bliver stængelassimilant. Stængelsukkulens med store mængder vandvæv i marven ledsages ofte af micro-phylli som hos Alluaudia og Euphorbia fimbriata. Andre Euphorbia (vortemælk) arter har fuldstændig opgivet blade ligesom kaktus. – Under emnet Rejseindtryk (Nordamerikanske ørkner) kan man se andre eksempler på microphylli og tab af blade i tørtiden som hos Idria og Fouquieria, mens Ephedra er rene stængelassimilanter.
En række planter fra varme klimater har vist sig at have to slags grønkorn, hvor den afvigende type er placeret i et cellelag kaldet strengskeden placeret omkring ledningsstrengene. Disse grønkorn er større og har tyndere lamelstakke (grana) med chlorofyl. Samtidig er strengskedecellerne tykvæggede og isolerede mod det øvrige bladkød af en korklamel (rød streg på fig. t.h.). Det har vist sig, at denne bladanatomi er koblet med et særligt vandbesparende fotosyntese-system kaldet C4, mens det normale kaldes C3. Navnet kommer af det første produkt, der dannes, når CO2 fixeres, nemlig en kulstofforbindelse med henholdsvis 4 og 3 kulstofatomer.
Majs (strengskedecelle vist herover) er en C4 plante, og dens fotosynteserate bliver ved med at stige ved højere lysintensitet i modsætning til C3 planter, der når et maksimum, hvor de er lysmættede og ikke kan øge fotosyntesen yderligere som vist på skemaet øverst t.h. Her er der også en simplificeret fremstilling af den normale C3-fotosyntese. Lysprocesserne i grana producerer energien (ATP og NADPH2) til at drive CO2 assimilationen. Det hvide korn er produceret stivelse. Det fremgår, at der under respirationsprocesserne slipper CO2 ud gennem stomata, og med stigende lysintensitet og temperatur udlignes CO2 assimilationen, så fotosynteseraten flader ud. På det nederste diagram er fotosyntesens forløb hos en C4 plante tilføjet. Det væsentlige er, at den ved respirationen producerede CO2 ikke slipper ud i det fri, men eksporteres til cellerne uden for strengskeden, hvor det gen-assimileres. Derfor kan fotosynteseraten blive ved med at stige med højere lysintensitet, men det betyder også en stejlere koncentrationsgradient for CO2 fra luften og ind i bladet. Derved kan åbningsraden af stomata mindskes, og det betyder en større modstand for den stomatære transpiration, og C4 fotosyntese er dermed vandbesparende.
Der findes ca. 8.100 C4 planter, hvoraf langt de fleste er énkimbladede urter, mens meget få er træer (Paulownia er et eksempel). I Danmark har vi kun én naturlig C4 plante. Det er sodaurt, Salsola cali, og den vokser på stranden ved foden af klitterne, hvor man ofte har den højeste lysintensitet og temperatur. Dens blade er sukkulente med et lag palisadeceller og derunder en strengskede, der dog ikke er fuldkommen, men afbrudt af vandvævet, der flere steder når ud til epidermis.
Sporobolus rigens er et C4 græs endemisk for Argentina og Chile. Det er endnu et eksempel på, at strengskeden kan være afbrudt. Den ses her som mørktfarvede celler ind mod det kompakte assimilationsvæv. Af andre xeromorfe karakterer bemærkes det centrale vandvæv, og styrkevævet omkring ledningstrengene fortsætter i en strengskede-forlængelse ud til epidermis. – Der er i Mellemøstens ørkner fundet nogle få Chenopodiaceer, der er i stand til at udføre C4 fotosyntese uden strengskede, idet to nødvendige enzym systemer holdes adskilt i henholdsvis cytoplasmaet og central vakuolen.
Der findes endnu et fotosyntesesystem, CAM-fotosyntese, der er en fysiologisk tilpasning til tilværelsen som xerofyt. Navnet kommer af, at systemet først blev fundet hos stenurtfamilien, Crassulaceae (Crassulacean Acid Metabolism), men nu kendes det fra mange andre familier som f.eks. Orchidaceae, Cactaceae og Bromeliaceae, men næsten kun hos sukkulente planter med meget vandvæv. Metoden går ud på, at CO2 i første omgang fixeres i oxaleddikesyre v.h.j. enzymet PEP (fosfoenol-pyrodruesyre) og derefter oplagres som æblesyre i central vakuolen. Det sker om natten, hvor luftfugtigheden er relativt høj, så transpirationen er lille selv om stomata er åbne. Om dagen, når lyset sætter gang i fotosyntesens lysprocesser, frigøres den oplagrede CO2 og assimileres som ved normal C3-fotosyntese, men med lukkede stomata, og det sparer på vandet. Planten t.h. er Crassula arta fra Sydafrika og Namibia.
Kort resumé af xeromorfe karakterer
Stor absorberende rodoverflade, proteoide rødder – top/rod forhold lille. Mikrofylli – ericoide og ind- eller tilbagerullede blade – stængelassimilation. Kraftig kutinisering af epidermis, lamellær kutikula nedsætter den kutikulære transpiration. Meget epikutikulært voks reflekterer stråling og reducerer bladets opvarmning. Beklædning med døde luftfyldte dækhår øger grænselaget og mindsker opvarmning. Vandabsorberende hår der opsuger regn og dug. Æteriske olier i kirtelhår og sekrethuler mindsker vands overfladespænding og trænger vandfasen tilbage i cellevæggen. Indsænkede spalteåbninger – mange/areal – lukning af overflødige stomata. Spalteåbninger i furer eller huler. Vandvæv i cortex, marv, hypodermis, mesofyl (garvestof-vakuoler). Veludviklet ledningsvæv og styrkevæv i hele planten og strengskedeforlængelser forhindrer sammenklapning. Kompakt assimilationsvæv og reducering af intercellulærrum mindsker det eksponerede vægareal C4-fotyntese og CAM-fotosyntese. Nogle planter tåler dehydrering bedre end andre – gælder især laver og mosser. Énårige planter klarer tørtiden som kim omgivet af en beskyttende frøskal.