Rundbladet soldug er den mest udbredte soldug art i Danmark. Den er cirkumpolar inklusive det sydlige Grønland. Den trives fint med konkurrence fra tørvemos (Sphagnum). På billedet th. vokser soldug tæt rundt om en grundvands regulere klitlavning i den rødbrune bræmme, hvor der hverken er for vådt eller tørt.
Drosera rotundifolia blomstrer i august. De 5-tallige blomster er kun åbne midt på dagen, og de sidder højt hævet over fælderne. I tørt og solrigt vejr kan slimen på tentaklerne tørre ud. På billedet herunder ses i midten et blad foldet sammen om et bytte som resultat af den på hovedsiden omtalte auxin regulerede vækstbevægelse.
Langbladet soldug - Drosera anglica
Langbladet soldug har i Danmark kun få voksesteder på højmoser og i fugtige klitheder i Jylland. Dens totale udbredelse er næsten cirkumpolar. Den findes dog ikke i Island og Grønland, men i Nordamerika når den op i Alaska. Der findes også nogle subtropiske forekomster på Hawaii og i det sydlige Europa og sydlige Japan. Den trives fint i sure omgivelser mellem Sphagnum. Den overvintrer ved vinterknopper kaldet hibernakler. Arten er selvbestøende og menes opstået som en diploid hybrid mellem den nordamerikanske D. linearis og D. rotundifolia. Drosera anglica x obovata er en steril hybrid med D. rotundifolia. Hybriden forveksles undertiden med Drosera intermedia.
Liden soldug - Drosera intermedia
Liden soldug findes især i moser, hvor den står under vådere forhold end de to øvrige danske arter, rundbladet og langbladet soldug. Den har hovedudbredelse i det vestlige Europa og østlige Nordamerika, men når ned i det nordlige Sydamerika. I tempereret klima overvintrer den ved hibernakler. Bladene er oprette, og formen er intermediær mellem de to førnævnte arter. Blomsterstanden virker sidestillet, idet den er bøjet til siden, mens den står opret centralt i rosetten hos de to andre danske arter. Bomstrer fra juni til august. På billedet t.h. vokser den sammen med blærerod Utricularia australis. – Drosera anglica x obovata er en steril hybrid med D. rotundifolia. Hybriden forveksles undertiden med Drosera intermedia.
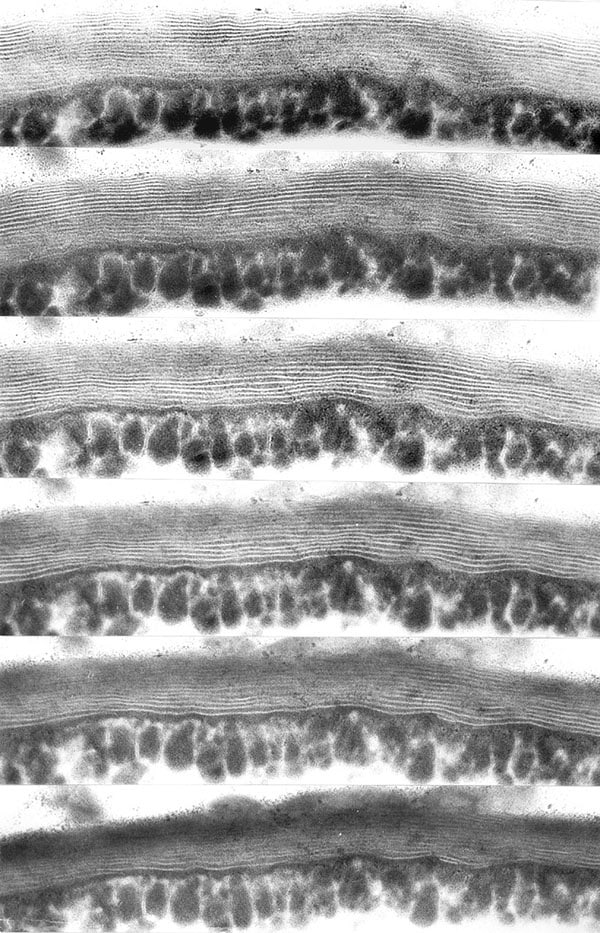
Hakea drupacea (= H. suaveolens). Developing polylamellate cuticle proper seen in a goniometer series with a 5° tilt (in same direction) from micrograph to micrograph. Photographed at 29.000x.
Heide-Jørgensen H S. 1981. Morphology and occurrence of different types of cuticular membranes. - In J Wattendorff and KJ Lendzian (eds.): The protective layers in higher plants and their function: Cuticular membranes and periderms. Conf. Abstr. Univ. Freiburg 1981: 10.
Abstract/Summary: Cuticles, in the widest sense of the word, are found all over the surface of higher plants but attain in thickness and complexity their most advanced development on aerial primary stems, leaves and fruits. On a genetic basis the morphology, including the ultrastructure of the cuticular complex, varies between species as well as within species and even within the same leaf depending on the epidermal cell type in question. However, some variation due to environmental influence may also be found.
Generally, going out from the inside, the outer epidermal wall consists of a layered wall of polysaccharides, a pectic zone, the cuticular layer, the cuticle proper, and an epicuticular wax layer. Ultrastructurally the cuticle proper may be one of four types. a) The lamellate type, composed of almost equally thick alternating cutin and wax lamellae; this type may be subdivided in relation to the extension of the lamellae. b) The dendritic type, where, more or less perpendicular to the surface, thin branching strands of electron dense material traverse an otherwise homogeneous and electron translucent matrix. c) The reticulate type, where electron dense material appears in a reticulum embedded in a translucent matrix. d) The homogeneous type, which appears more or less electron dense.
The cuticular layer is even more variable. It may be absent or it may form the bulk of the outer wall. Ultrastructurally it consists of an anastomosed network of electron dense fibrillae, interpreted as pectins, between which cutin and wax are deposited in highly variable quantities.
At the present state of knowledge it is not clear if this classification of cuticles should be further graduated, or if the number of types should be reduced since some of the types may be related only to certain steps during cell wall differentiation, and some may merely be artefacts due to incomplete fixation. However, considering the wide spectrum of functions and the diversity of environmental conditions it is readily acceptable that structurally different cuticle types have developed.