Saprophytes grow on and get nourishment from dead organic matter, and in the past a number of achlorophyllous plants without photosynthesis were classified as saprophytes, but here botanists were wrong. It has been shown that all these plants in fact get their nutrition from other plants with photosynthesis via a mycorrhizal fungus as an intermediary. Only fungi can directly utilize dead organic matter, so they can still be called saprophytes. Plants that obtain nutrition from other plants via mycorrhiza are called mycoheterotrophic plants, while parasitic plants (parasites) are plants that obtain water and nutrition from their host plants via a modified root called a haustorium. Both among parasitic plants and mycoheterotrophic plants, there are species that completely lack photosynthesis, while others themselves produce at least some carbon compounds via photosynthesis, and some plants are only mycoheterotrophic at certain stages of their life cycle, such as prothalli (gametophytes) in some ferns and seeds in some orchids. If the plant receives both nutrition through photosynthesis and mycoheterotrophy, it is called a mixotroph.
Mycorrhiza is a crucial element in mycoheterotrophic plants. There are several forms of mycorrhiza, the two most wide-spread being ectotrophic mycorrhiza formed by fungi with aboveground fruiting bodies (toadstools, Basidiomycetes) and endotrophic mycorrhiza also called VA mycorrhiza formed by underground fungi (Zygomycetes) without aboveground fruiting bodies. Above, ectotrophic mycorrhiza can be seen on the roots of a beech (Fagus silvatica). The fungus-infected roots apparently swell, as the hyphae of the fungus form a dense sheath around the root. Some hyphae grow in between the cells, where nutrients are exchanged across the cell wall and cell membrane. At left two different fungal species are involved, one with light, the other with dark hyphae. Note on the left side of the picture a thin, uninfected lateral root. Photo at left is from Naturens Verden.
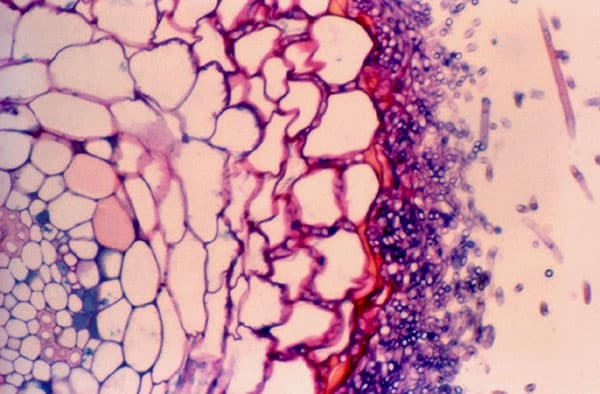
In VA mycorrhiza, no sheath is formed, but the hyphae grow directly into the host’s cells, however without breaking through the cell membrane. Inside the cells, the hyphae form arbuscules, also called hyphal keys with reference to yarn keys. The arbuscules are consumed with age by the host, which benefits from the nourishment. In addition, some hyphae form vesicles at the tip, which act as the fungus’s storage cells for lipids (round in the picture at right), which it receives from the host. Photo: Naturens Verden.
The first mycoheterotrophic flowering plant was recognized in 1889, but since then the number has steadily increased and from 1994 to 2013 alone, the number increased from 413 to 517, and the number in 2022 is probably in the region of 600 spread over more than 80 genera. One reason for the uncertainty is that, just as you cannot immediately tell whether a green plant is a hemiparasite with the haustoria sitting on roots underground, you also cannot see whether a green plant or a chlorophyll-free plant is a mycoheterotroph without examining, whether it has mycorrhiza. In the following, a few examples of mycoheterotrophic plants are given, and if you want to see more examples, the following link, where there are also literature references, is recommended: https://parasiticplants.siu.edu/Mycotrophs/Mycotrophs.html
There is a mycoheterotrophic liverwort Aneura mirabilis (formerly Cryptothallus mirabilis). The species has been found in northern Europe and at a single location in Greenland. The plant is small and completely white, as it has no chlorophyll, and it lives under moss cover or in peat soil. It is dioecious with female and male plants, where the female plant is up to 3 cm and significantly larger than the male plant. The picture at right shows a male plant with antheridia. Photo: Michael Lüth.
Most ferns have an above-ground green prothalus with photosynthesis, but i.a. in the snake-tongue family, Ophio-glossaceae, and in clubmosses, the prothallus (gametophyte) is subterranean and without chlorophyll, so at that stage of the plants’ life cycle they are mycoheterotrophs. The picture at left shows serpent’s Tongue, Ophioglossum vulgare.
Parasitaxus usta from New Caledonia is a gymnosperm that grows on Falcatifolium taxoides, both belonging to the Podocarpaceae. It has been much debated whether P. usta is a holoparasite or a mycoheterotrophic plant. Young shoots have small scale-shaped red leaves without photosynthesis. A haustorium with xylem to xylem contact has not been found, as in all parasitic plants, but transfer of 13C via VA mycorrhiza has been found. Everything points to a tripartite connection that can neither be considered a holoparasite nor a mycoheterotroph. As a rule, P. usta shoots from the host’s roots, while the shoots from the host’s trunk up to 1.20 m above the ground are a rarity. Photo: B Suprin.
There are a large number of monocot mycoheterotrophic species in the tropics and subtropics from the families Burmanniaceae, Corsiaceae, Iridaceae, Petrosaviaceae, Triuridaceae and Orchidaceae, but only the latter family is represented in Denmark with three species.
Bird’s nest orchid, Neottia nidus-avis, photographed on Møns Klint is seen at left. It has short tangled roots that look like a bird’s nest. The species is widespread in most of Europe and reaches east to the Caucasus and south to North Africa. As it is independent of photosynthesis, it can grow in deep shade, often in beech forests on calcareous subsoil. The mycorrhizal fungus belongs to the genus Sebacina. The plant can grow 30 cm high and blooms in June-July. Pollination is done by flies and ants, but self-pollination can occur.
Coral root, Corallorhiza trifida, is also found in Denmark. It is widespread in Eurasia and North America. In Greenland it is found on the west coast up to Disko. The plant can be seen at left. It has branched rhizomes reminiscent of corals, but no actual roots. The stem is yellow-green and contains a little chlorophyll, but photosynthesis is insufficient, so it also receives nourishment from fungi in the genus Tomentella, which have willow and birch mycorrhiza. Coral root flowers in May-June and the lip is white with red dots. Several years can pass between flowering. – The three other photos show Corallorhiza maculata, which is completely devoid of chlorophyll. It is widespread in North America and exists in three varieties that use mushrooms from the family Russulaceae. It stays away from the lowlands and is found especially in mountain forests. The native people have used dried stems to brew tea and use them as a remedy for colds, pneumonia and skin irritation.
The 3rd mycoheterotrophic orchid in Denmark is Spurred coral-root, Epipogium apyllum, which is very rare with long periods when it is only found underground. Last seen blooming in 2006 and 2021. The name comes from a crack on the lip that faces upwards with the spur. The stem is hollow. The species is found in Europe and northern Asia. There are three other species in the genus with primarily tropical and subtropical distribution. The drawing of E. aphyllum at left is from C.A.M. Lindman 1973: Nordens flora.
There are many other mycoheterotrophic orchids in the genera Cymbidium, Eulophia, Gastrodia, Cephalanthera, Epipactis, Platanthera, Rhizantella and many more. Most of the genera mentioned have many species, but only one or a few are mycoheterotrophic or mixotrophic.
A further three orchid genera represented in Denmark have mycoheterotrophic species. This applies to the Cephalanthera, where C. austinae from North America is completely devoid of chlorophyll. In Japan, there is a butterfly orchid, Platanthera minor with ectomycorrhiza and mixotrophic lifestyle, and the Denmark-growing broad hellebore, Epipactis helleborine (right photo), which was introduced to the USA at the end of the 19th century has now spread widely in North America, where individuals have been found that are not only mycoheterotrophic in the germ stage, but remain so and thus lack photosynthesis in the adult stage.
Rhizanthella gardneri (left) is an orchid that was first discovered in 1928 by a farmer in SW Australia when burst flowers suddenly erupted from the ground while he was plowing his field. The plant grows and blooms completely underground. It has no chlorophyll and gets its nourishment through fungi that have mycorrhiza with green autotrophic plants. The orchid is thus a mycoheterotrophic plant. The approx. 150 tightly packed small flowers attract termites as pollinators, and they are so strongly scented that humans can pick up the scent through the soil. Only six populations are known, so the species is considered to be in great danger of extinction. Photo: Jean & Fred, Flickr.
The following examples come from the heather family, Ericaceae, in the broadest sense, as several previously recognized families are now incorporated into Ericaceae. In Denmark, yellow bird’s-nest, Monotropa hypopitys (above) is a mycoheterotroph and completely without photosynthesis. Genetic studies indicate that the species will henceforth be in its own genus and be called Hypopitys monotropa. The perennial plant is widespread in the temperate zone of the northern hemisphere. The inflorescences develop from the roots, and the plant have no actual stems. The 5-numbered flowers open in Denmark from June to August. The flowers are drooping at first, but straighten during the ripening of the capsules. There are several subspecies, some prefer deciduous forest others coniferous forest.
Monotropa uniflora, Indian pipe, is found in temperate North America and Asia as well as isolated in northern South America. It is a perennial plant where the inflorescences develop quickly but only last a few days. There are varieties where the plant is pink or red. The photos above are from Vancouver Island. The associated mycorrhizal fungi are from the Russulaceae. Pollination takes place with flies and bumblebees, and the inflorescence is straightened in the fruiting stage as in M. hypopitys.
Allotropa virgate, sugarstick, is a perennial plant up to 50 cm tall with a rhizome and distribution in western North America, where in mountain forests it goes up to 3,000 m. The picture at left is from Vancouver Island and shows a growing inflorescence. The stem gradually becomes red-striped, and the leaves, as in other mycoheterotrophic plants, are reduced to scales without photosynthesis. Tricholoma matsutake and Tricholoma magnivelare are indicated as the mycorrhizal fungi used. The flowers often lack sepals. There are 5 free white petals forming a cup-shaped flower with red anthers. The flower contains nectar.
The wintergreen family Pyrolaceae is also included in Ericaceae, and here there are mixotrophic species in all four genera represented in Denmark. The species above is Pyrola picta ssp. aphylla, which is endemic to California. It is leafless and mycoheterotrophic throughout its life, while P. picta and the other species in the genus are considered mixotrophic. This also applies to the wintergreen species in the genera Chimaphila, Moneses and Orthilia.
Pterospora andromeda, pinedrops, is the only species in the genus. It is found in both eastern and western North America and reaches down into northern Mexico. It prefers coniferous forest. The inflorescence can be 1 m high. The photos above are from the San Bernardino Mountains in California and show mature fruit stands where the 5-compartment capsules are wooded. It blooms from June-August, and the flowers are hanging and bell-shaped with red sepals and a whitish crown. There seems to be a difference as to which fungus it has symbiosis with in the western and eastern parts of the distribution, such that Rhizopogon salebrosus is used in the west and Rhizopogon kretzerae in the east. Pterospora is considered closely related to the following species Sarcodes sanguinea.
Sarcodes sanguinea, snowplant, is another monotypic mycoheterotrophic species from North America with scattered occurrence in Oregon, California and Nevada. The plant can grow up to 30 cm high. The genus name alludes to the fact that the inflorescence is meaty. It is also glandular. It blooms in early spring, when there may still be some snow in the mountains. Regarding fungi it is highly host-specific and can only form a partnership with the basidiomycete Rhizopogon ellenae, which forms ectomycorrhizae. The two photos at right is by Joe Antos.
Herb paris, Paris quadrifolia
This was briefly about mycoheterotrophic plants and mainly those that I myself have been around. The name is a bit difficult, but the plants can’t help it – they’re just interesting.
It has recently been shown that far more of the common green forest floor photosynthetic plants, and probably even the vast majority, are partially nourished via mycorrhizal fungi, which in turn obtain carbon compounds from their hosts. Such a mutually beneficial relationship is called symbiosis. It has been proven i.a. in several horsetails and ferns and flowering plants such as wood anemone (Anemone nemorosa), herb paris (Paris quadrifolia), wood sorrel (Oxalis acetosella), and wood cranesbill (Geranium sylvaticum). One can argue that these plants are parasitic plants/parasites (on the fungus), but since it is structurally and physiologically different from the haustorial parasites, I prefer to use the term mycoheterotrophic plants, even if it is a difficult name.
H. S. Heide-Jørgensen, February 2022, translated March 2024.